

Respirasome Proteins Are Regulated by Sex-Hormone Interactions in the Brain
- PMID: 36499081
- PMCID: PMC9741126
- DOI: 10.3390/ijms232314754
Respirasome Proteins Are Regulated by Sex-Hormone Interactions in the Brain
Abstract
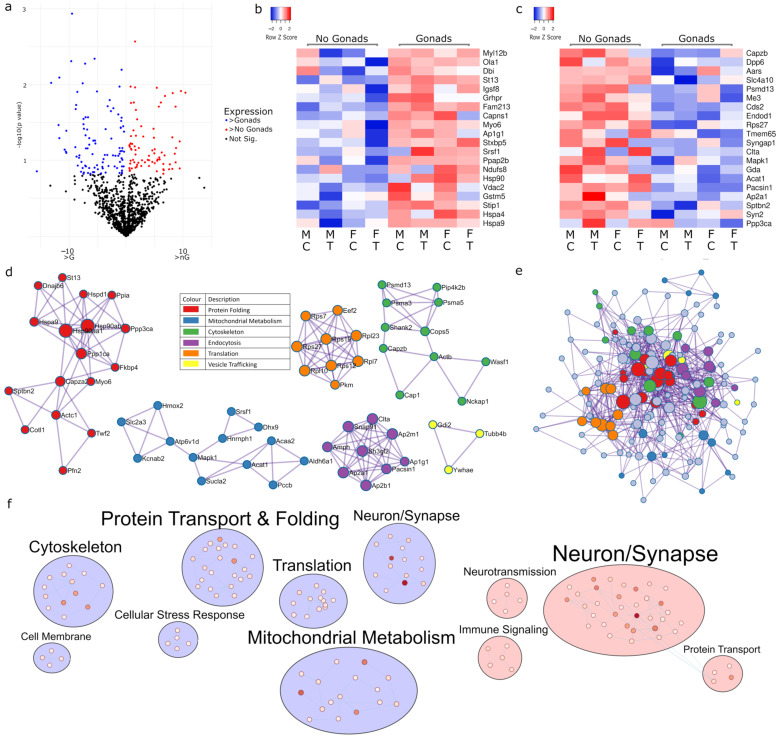
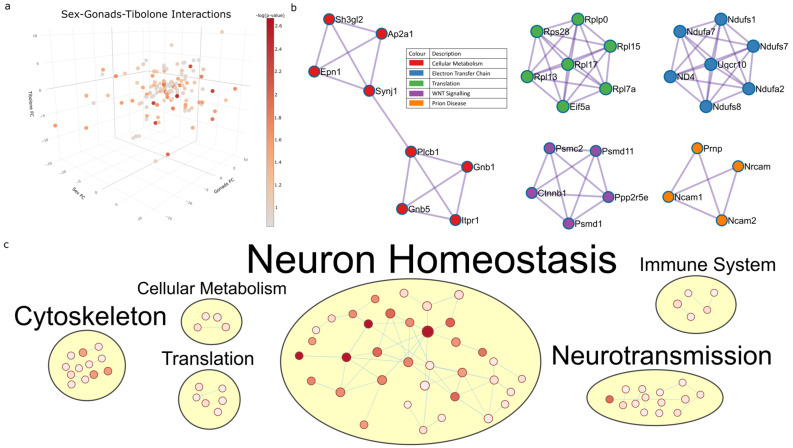
The existence of sex differences in disease incidence is attributed, in part, to sex differences in metabolism. Uncovering the precise mechanism driving these differences is an extraordinarily complex process influenced by genetics, endogenous hormones, sex-specific lifetime events, individual differences and external environmental/social factors. In fact, such differences may be subtle, but across a life span, increase susceptibility to a pathology. Whilst research persists in the hope of discovering an elegant biological mechanism to underpin sex differences in disease, here, we show, for the first time, that such a mechanism may be subtle in nature but influenced by multiple sex-specific factors. A proteomic dataset was generated from a gonadectomized mouse model treated with Tibolone, a menopausal hormone therapy. Following functional enrichment analysis, we identified that Alzheimer's disease and the electron transport chain-associated pathways were regulated by sex-hormone interactions. Specifically, we identified that the expression of three respirasome proteins, NDUFA2, NDUFA7 and UQCR10, is significantly altered by compounding factors that contribute to sex differences. These proteins function in bioenergetics and produce reactive oxygen species, which are each dysregulated in many diseases with sex differences in incidence. We show sex-specific reprogrammed responses to Tibolone following gonadectomy, which primarily influence the expression of proteins contributing to metabolic pathways. This further infers that metabolic differences may underpin the observed sex differences in disease, but also that hormone therapy research now has potential in exploring sex-specific interventions to produce an effective method of prevention or treatment.
Keywords: Tibolone; gonadectomy; mitochondria; proteome; respirasome; sex differences.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
-
- National Institutes of Health NIH guidelines on the inclusion of women and minorities as subjects in clinical research. Fed. Regist. 1994;59:1408–1413.
-
- Moisan F., Kab S., Mohamed F., Canonico M., Le Guern M., Quintin C., Carcaillon L., Nicolau J., Duport N., Singh-Manoux A. Parkinson disease male-to-female ratios increase with age: French nationwide study and meta-analysis. J. Neurol. Neurosurg. Psychiatry. 2016;87:952–957. doi: 10.1136/jnnp-2015-312283. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
