

Trigeminal Sensory Supply Is Essential for Motor Recovery after Facial Nerve Injury
- PMID: 36499425
- PMCID: PMC9740813
- DOI: 10.3390/ijms232315101
Trigeminal Sensory Supply Is Essential for Motor Recovery after Facial Nerve Injury
Abstract
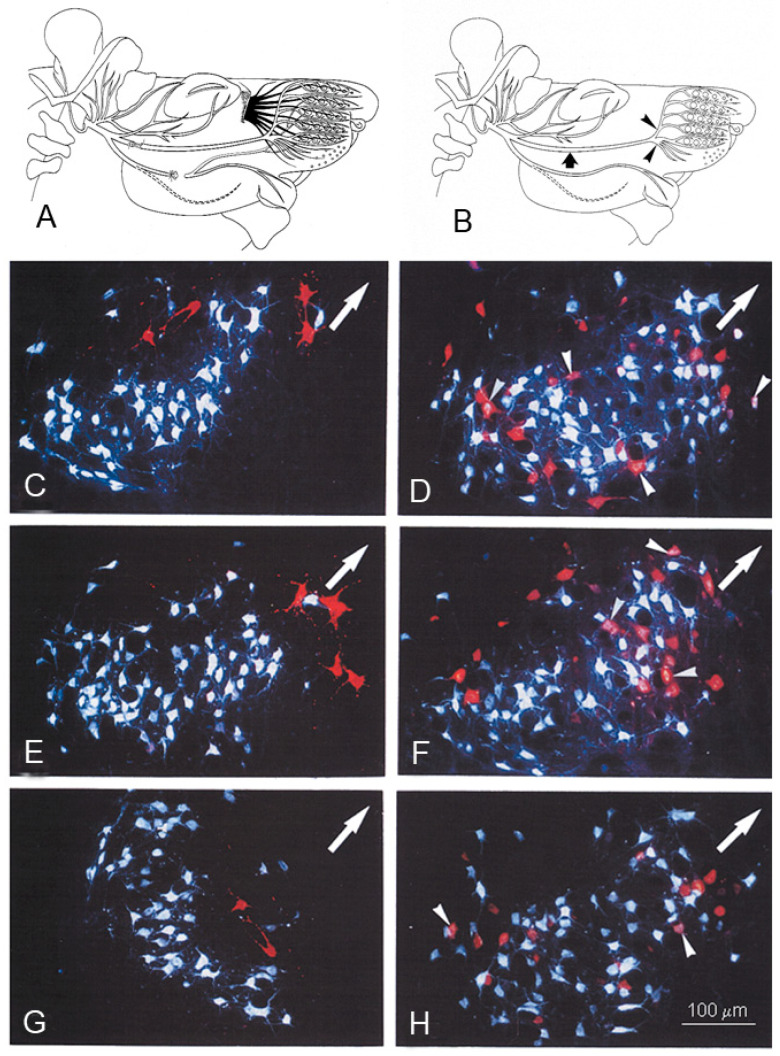
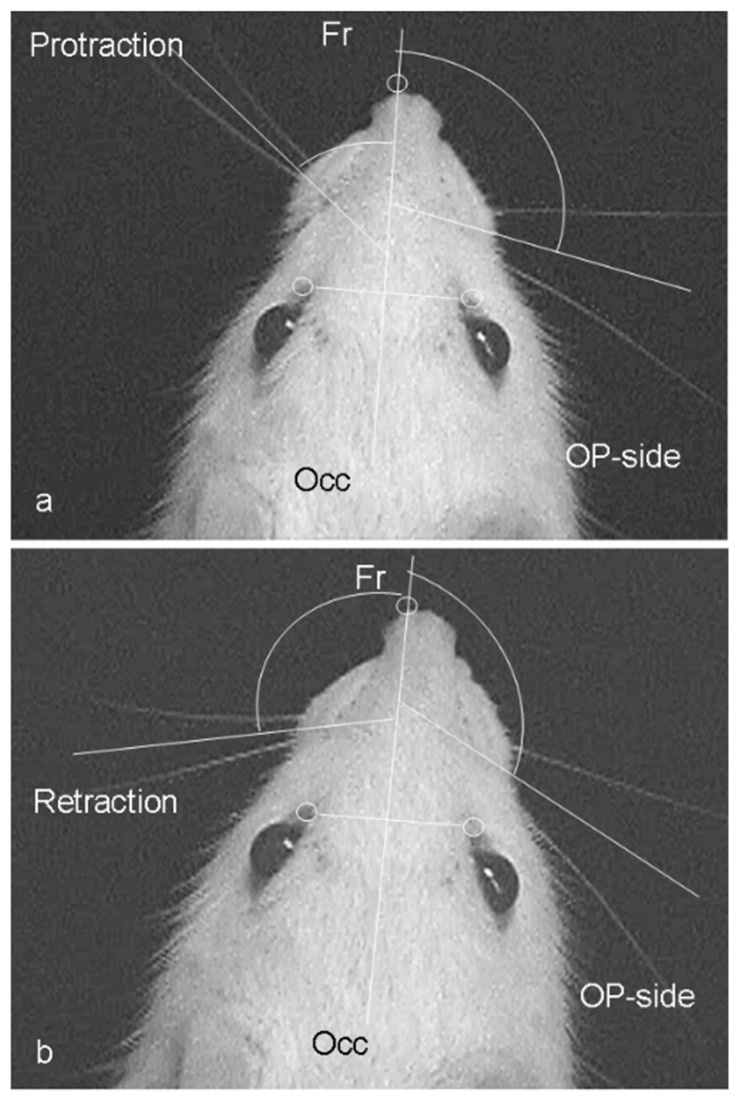
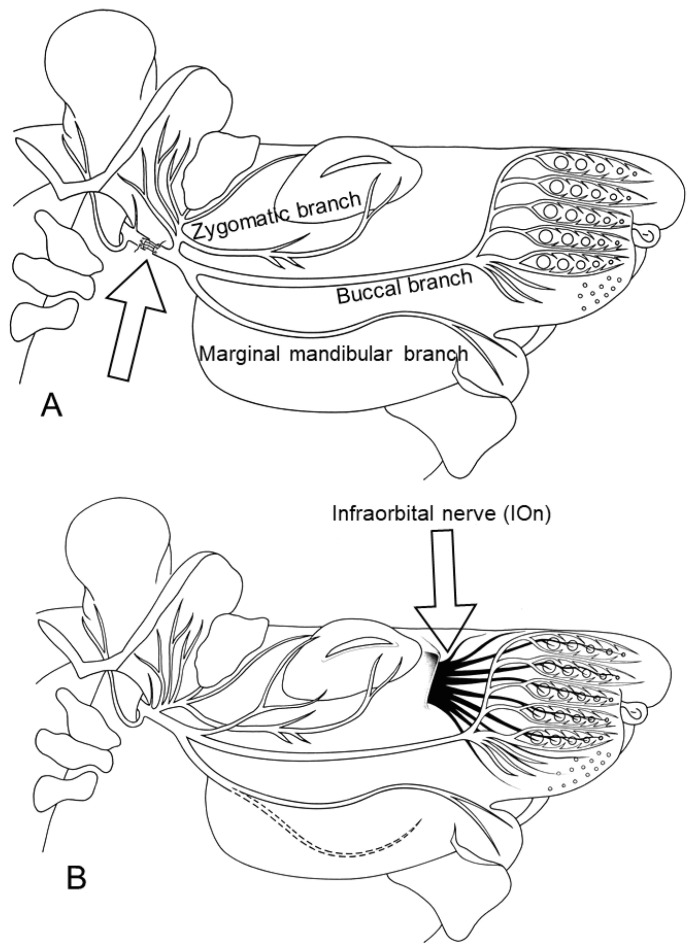
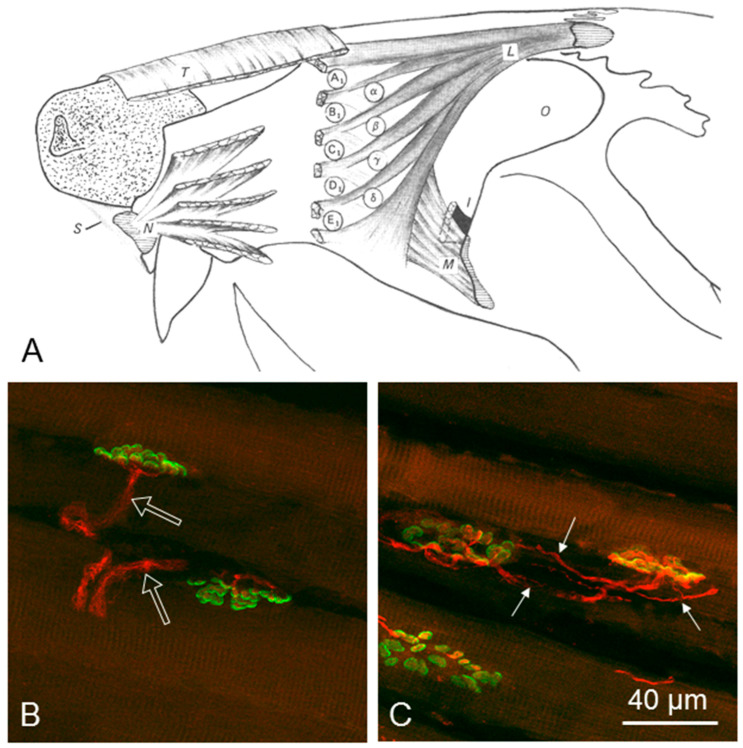
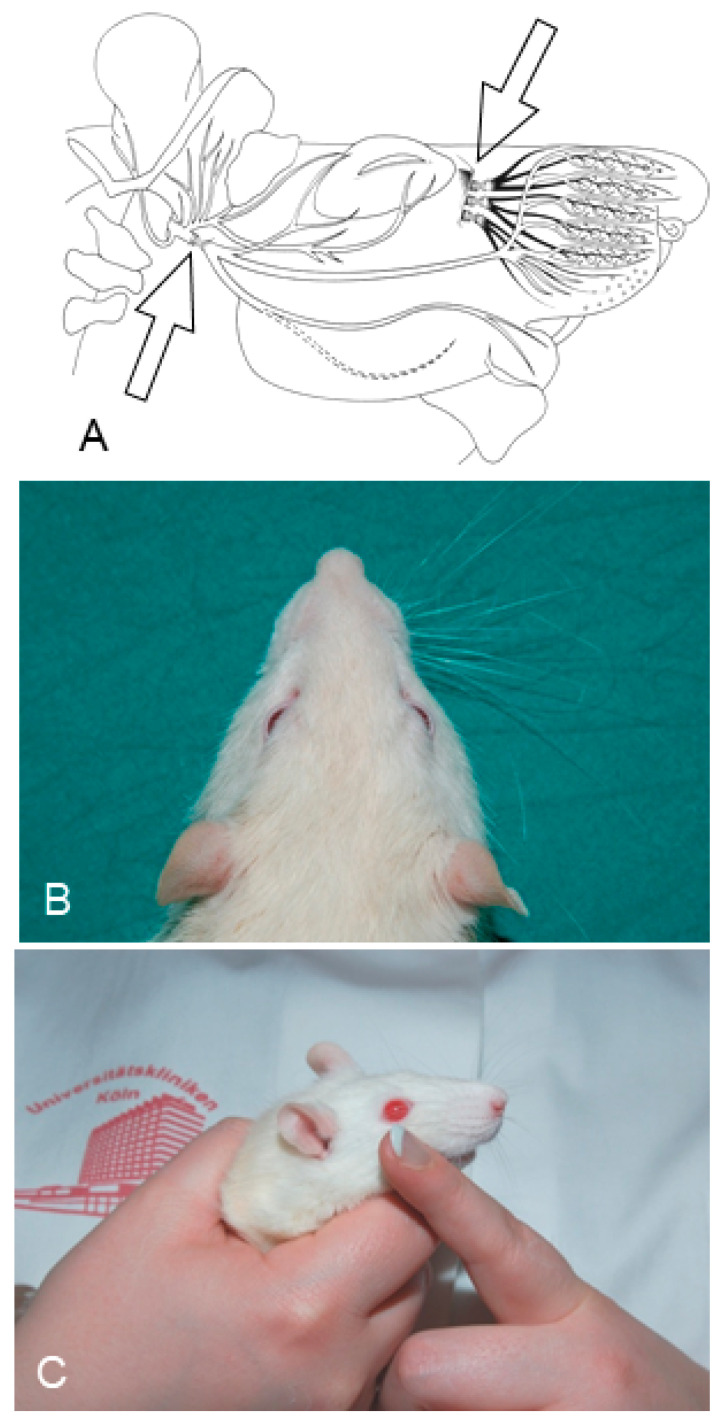
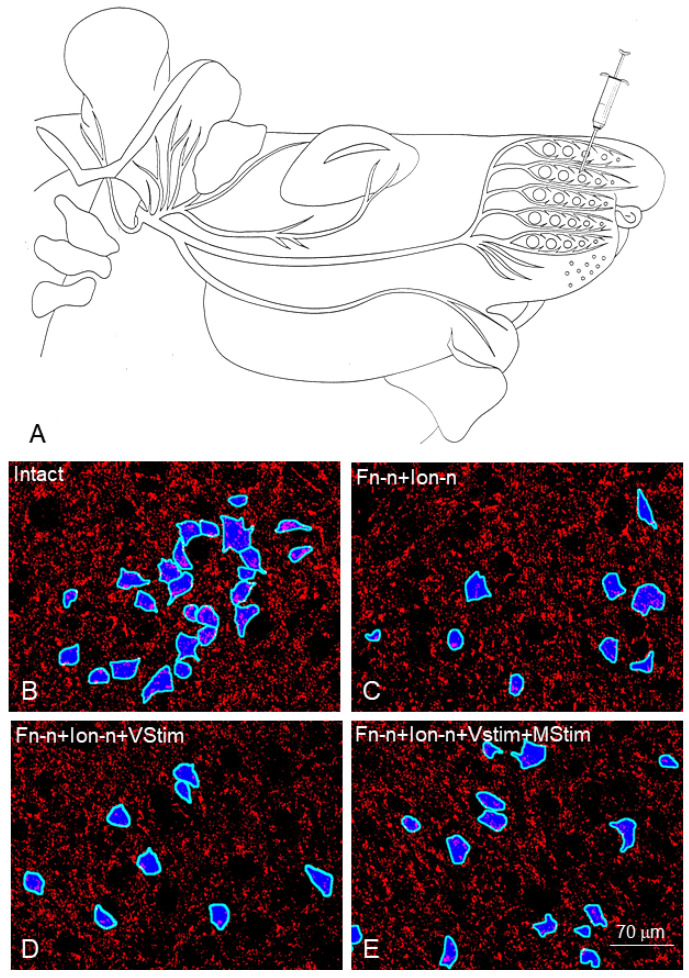
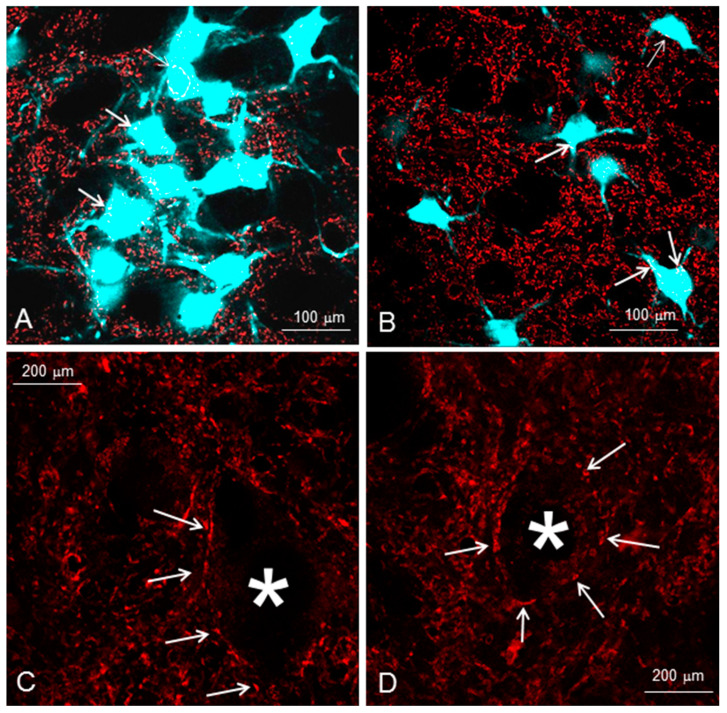
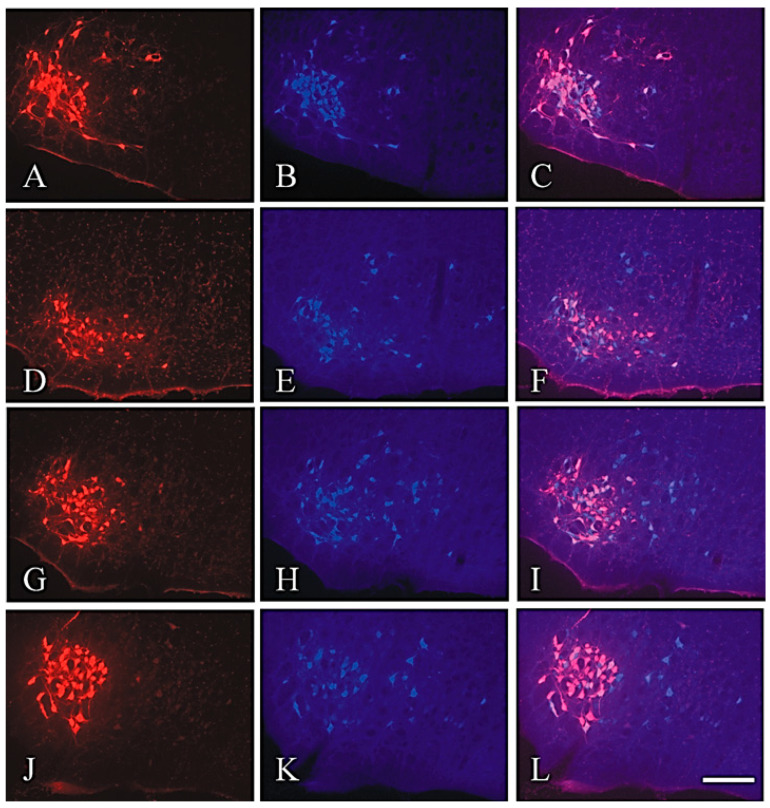
Recovery of mimic function after facial nerve transection is poor. The successful regrowth of regenerating motor nerve fibers to reinnervate their targets is compromised by (i) poor axonal navigation and excessive collateral branching, (ii) abnormal exchange of nerve impulses between adjacent regrowing axons, namely axonal crosstalk, and (iii) insufficient synaptic input to the axotomized facial motoneurons. As a result, axotomized motoneurons become hyperexcitable but unable to discharge. We review our findings, which have addressed the poor return of mimic function after facial nerve injuries, by testing the hypothesized detrimental component, and we propose that intensifying the trigeminal sensory input to axotomized and electrophysiologically silent facial motoneurons improves the specificity of the reinnervation of appropriate targets. We compared behavioral, functional, and morphological parameters after single reconstructive surgery of the facial nerve (or its buccal branch) with those obtained after identical facial nerve surgery, but combined with direct or indirect stimulation of the ipsilateral infraorbital nerve. We found that both methods of trigeminal sensory stimulation, i.e., stimulation of the vibrissal hairs and manual stimulation of the whisker pad, were beneficial for the outcome through improvement of the quality of target reinnervation and recovery of vibrissal motor performance.
Keywords: axotomy; facial nerve; morphological correlation; motion analysis; motoneuron; rat; recovery of function; stimulation; trigeminal nerve; vibrissal whisking.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Stimulation of trigeminal afferents improves motor recovery after facial nerve injury: functional, electrophysiological and morphological proofs.Adv Anat Embryol Cell Biol. 2013;213:1-105, vii. doi: 10.1007/978-3-642-33311-8_1. Adv Anat Embryol Cell Biol. 2013. PMID: 23322155
-
Putative roles of soluble trophic factors in facial nerve regeneration, target reinnervation, and recovery of vibrissal whisking.Exp Neurol. 2018 Feb;300:100-110. doi: 10.1016/j.expneurol.2017.10.029. Epub 2017 Nov 8. Exp Neurol. 2018. PMID: 29104116 Review.
-
Non-invasive stimulation of the vibrissal pad improves recovery of whisking function after simultaneous lesion of the facial and infraorbital nerves in rats.Exp Brain Res. 2011 Jul;212(1):65-79. doi: 10.1007/s00221-011-2697-9. Epub 2011 Apr 28. Exp Brain Res. 2011. PMID: 21526334
-
Contralateral trigeminal nerve lesion reduces polyneuronal muscle innervation after facial nerve repair in rats.Eur J Neurosci. 1999 Apr;11(4):1369-78. doi: 10.1046/j.1460-9568.1999.00545.x. Eur J Neurosci. 1999. PMID: 10103132
-
Intrinsic and therapeutic factors determining the recovery of motor function after peripheral nerve transection.Ann Anat. 2011 Jul;193(4):286-303. doi: 10.1016/j.aanat.2011.02.014. Epub 2011 Mar 12. Ann Anat. 2011. PMID: 21458252 Review.
Cited by
-
Central Facial Nervous System Biomolecules Involved in Peripheral Facial Nerve Injury Responses and Potential Therapeutic Strategies.Antioxidants (Basel). 2023 May 1;12(5):1036. doi: 10.3390/antiox12051036. Antioxidants (Basel). 2023. PMID: 37237902 Free PMC article. Review.
-
[Clinical analysis of transcranial facial nerve bridging with interpositional graft for the treatment of facial nerve injury].Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2024 May;38(5):376-379. doi: 10.13201/j.issn.2096-7993.2024.05.005. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2024. PMID: 38686472 Free PMC article. Chinese.
-
Neurological Complications following Surgical Treatments of the Lower Molars.Int J Dent. 2024 Sep 9;2024:5415597. doi: 10.1155/2024/5415597. eCollection 2024. Int J Dent. 2024. PMID: 39286455 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
