

Microbial detoxification of mycotoxins in food
- PMID: 36504774
- PMCID: PMC9726736
- DOI: 10.3389/fmicb.2022.957148
Microbial detoxification of mycotoxins in food
Abstract
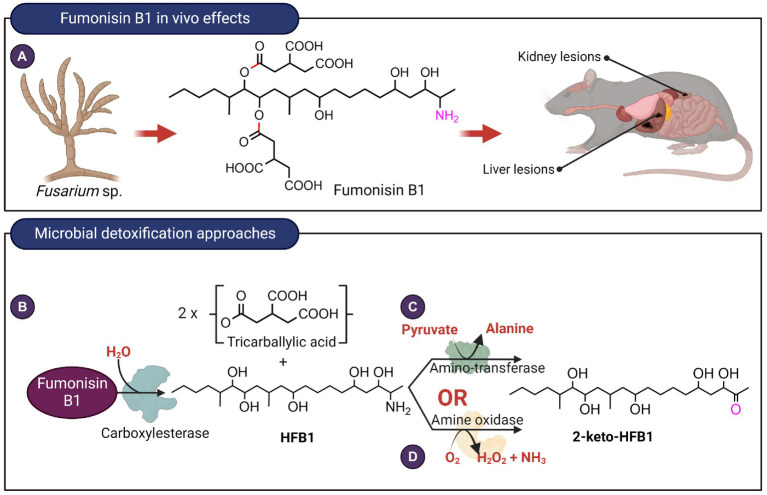
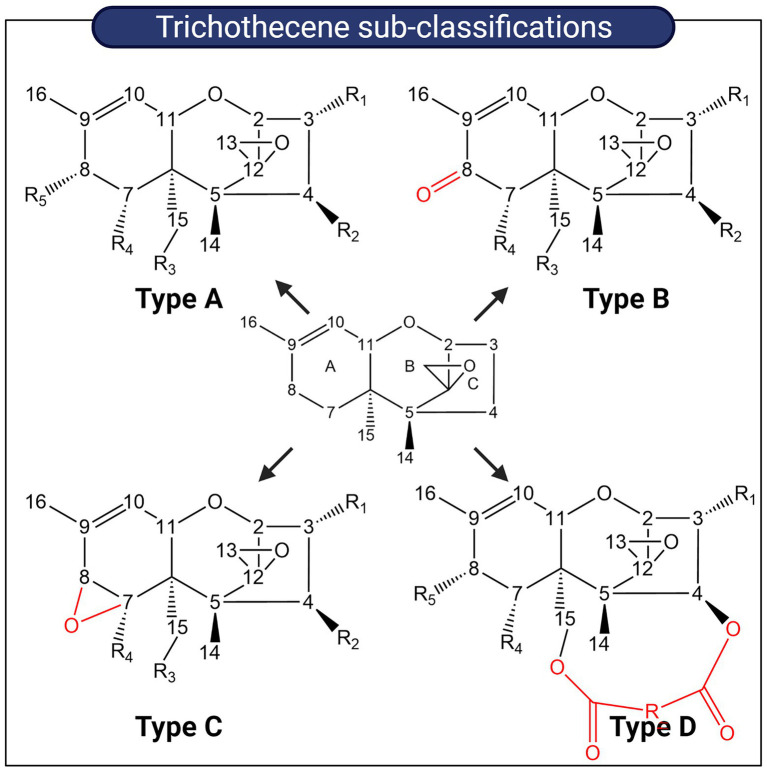
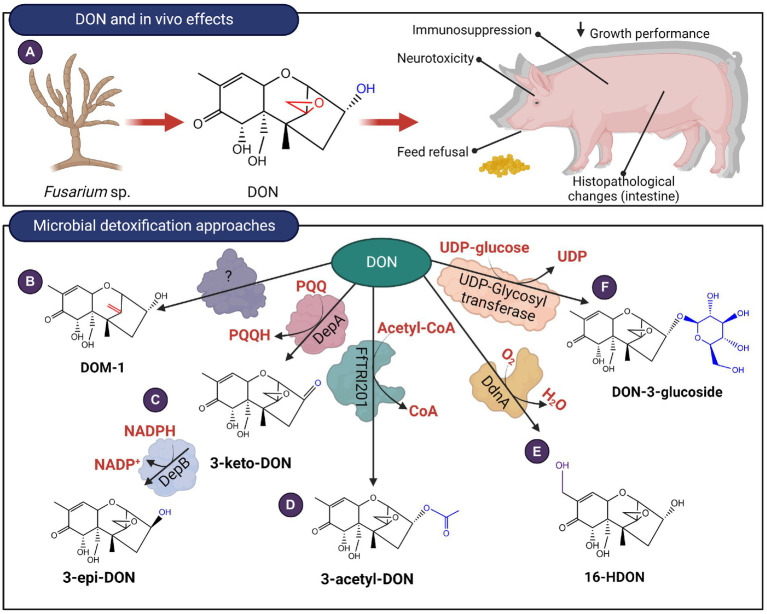
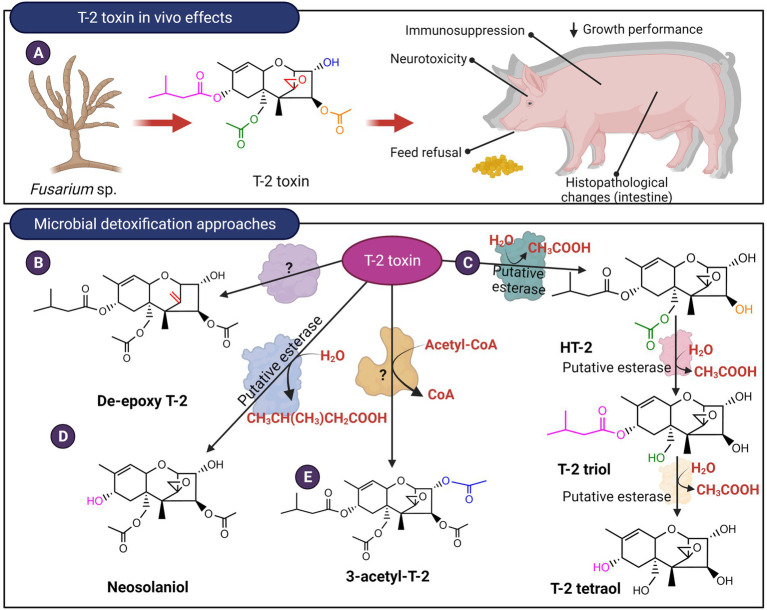
Mycotoxins are toxic secondary metabolites produced by certain genera of fungi including but not limited to Fusarium, Aspergillus, and Penicillium. Their persistence in agricultural commodities poses a significant food safety issue owing to their carcinogenic, teratogenic, and immunosuppressive effects. Due to their inherent stability, mycotoxin levels in contaminated food often exceed the prescribed regulatory thresholds posing a risk to both humans and livestock. Although physical and chemical methods have been applied to remove mycotoxins, these approaches may reduce the nutrient quality and organoleptic properties of food. Microbial transformation of mycotoxins is a promising alternative for mycotoxin detoxification as it is more specific and environmentally friendly compared to physical/chemical methods. Here we review the biological detoxification of the major mycotoxins with a focus on microbial enzymes.
Keywords: Mycotoxins; T2; aflatoxins; citrinin; deoxynivalenol; ochratoxin; patulin; zearaleneone.
Copyright © 2022 Abraham, Chan, Zhou and Seah.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
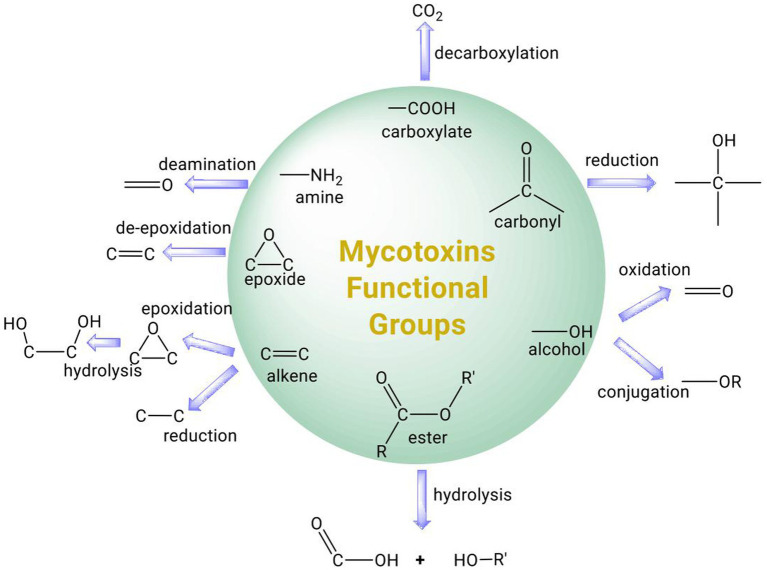
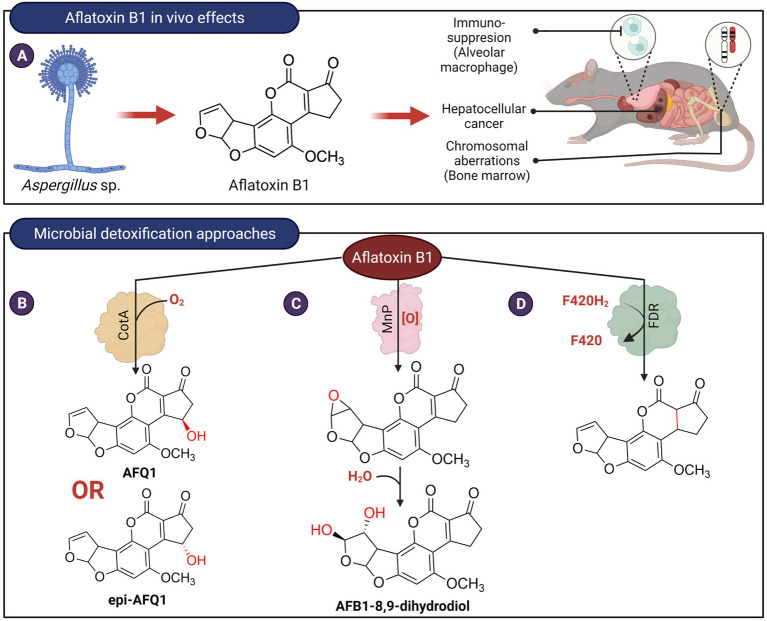
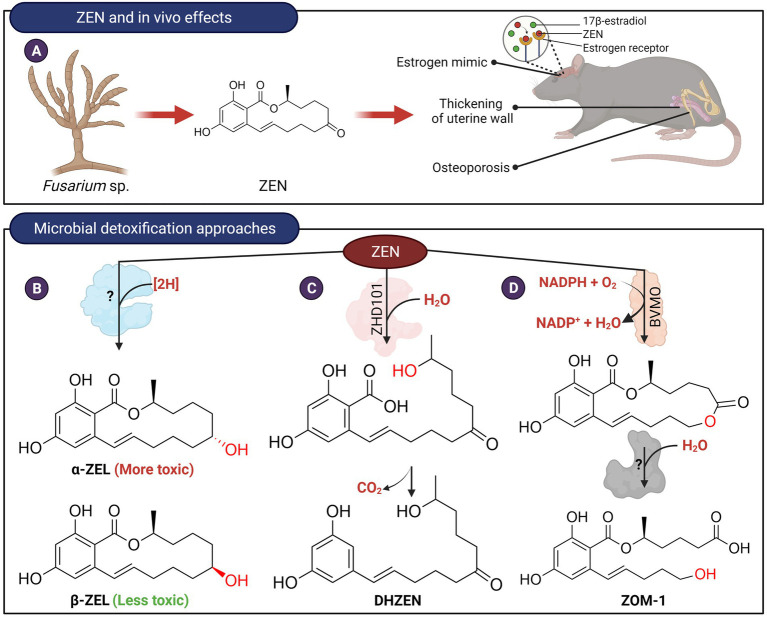
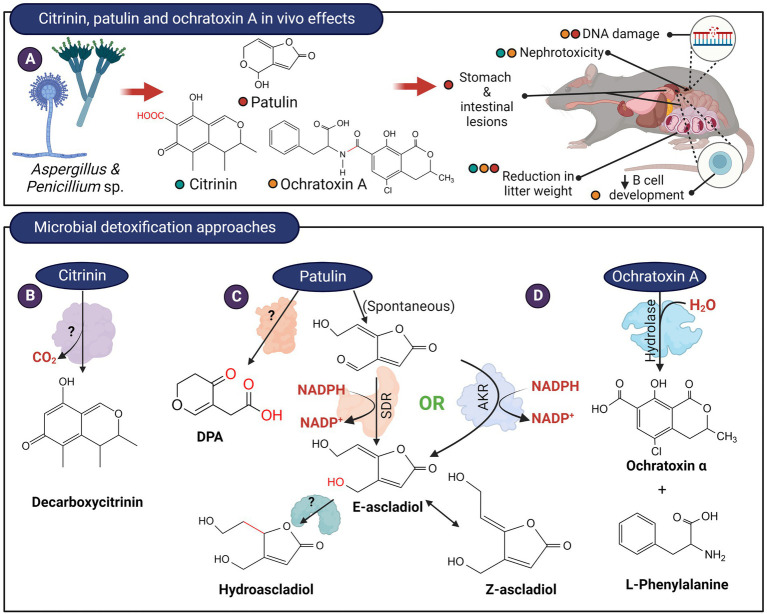
References
-
- Afriyie-Gyawu E., Wang Z., Ankrah N. A., Xu L., Johnson N. M., Tang L., et al. (2008). NovaSil clay does not affect the concentrations of vitamins a and E and nutrient minerals in serum samples from Ghanaians at high risk for aflatoxicosis. Food Addit. Contam. Part Chem. Anal. Control. Expo. Risk Assess. 25, 872–884. doi: 10.1080/02652030701854758, PMID: - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases