

RANKL-responsive epigenetic mechanism reprograms macrophages into bone-resorbing osteoclasts
- PMID: 36513810
- PMCID: PMC9794822
- DOI: 10.1038/s41423-022-00959-x
RANKL-responsive epigenetic mechanism reprograms macrophages into bone-resorbing osteoclasts
Abstract
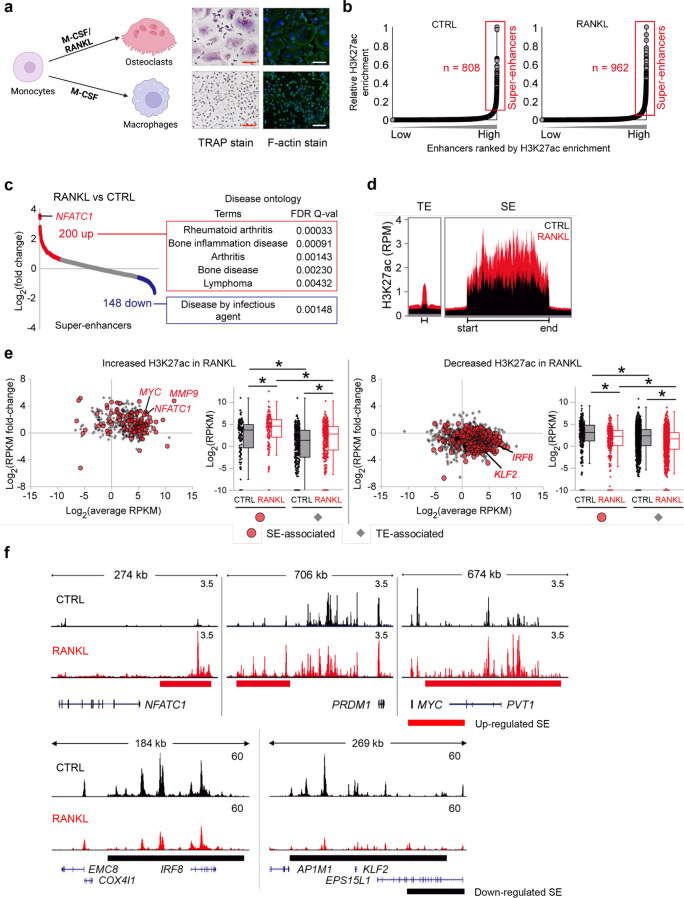
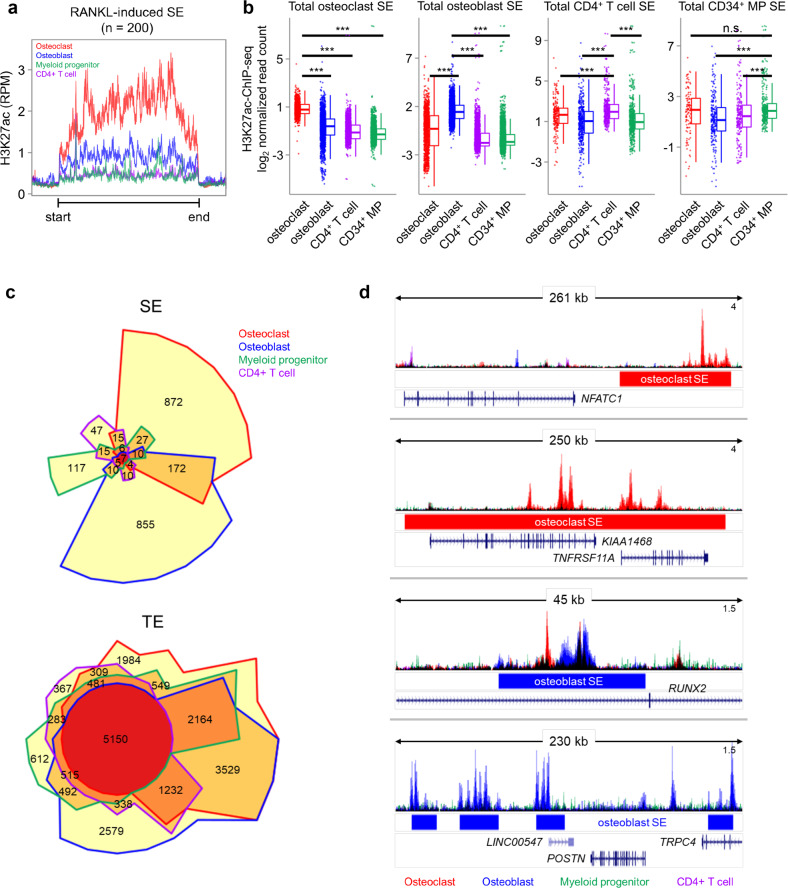
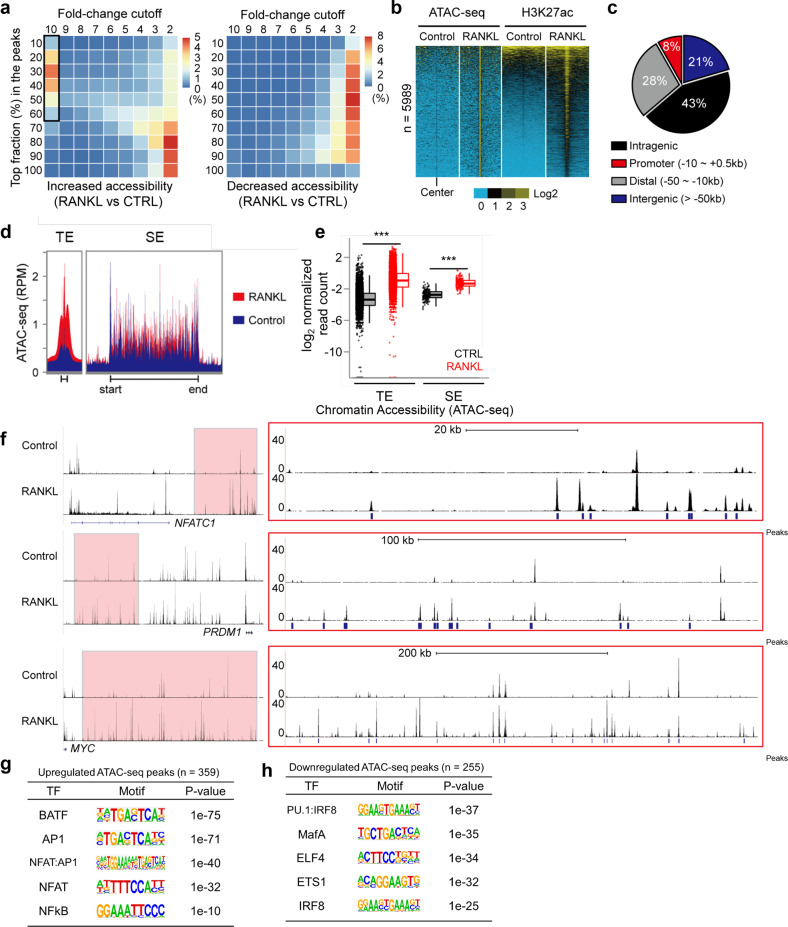
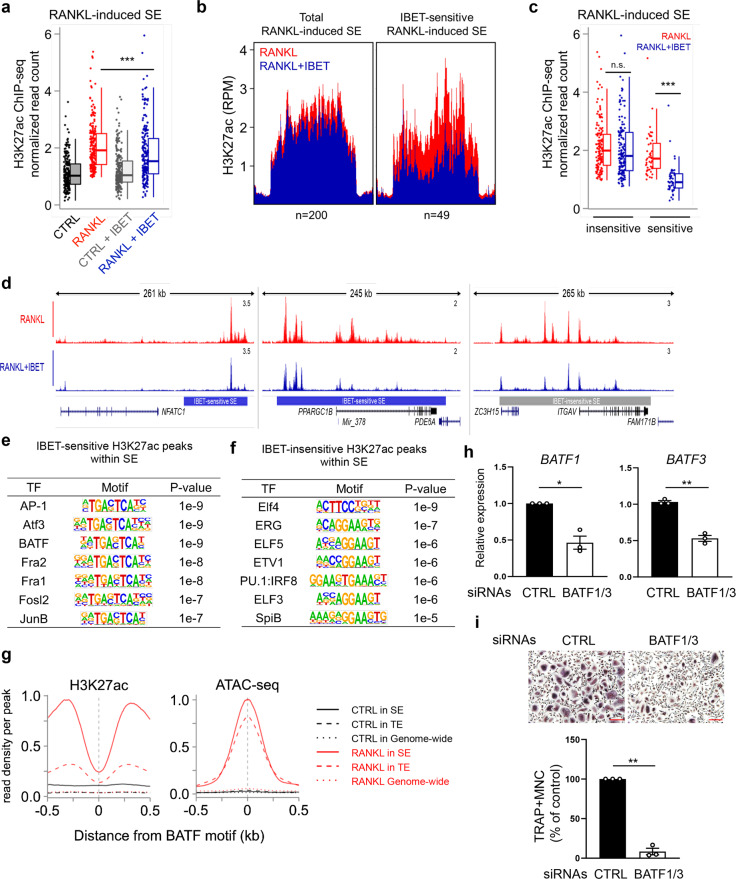
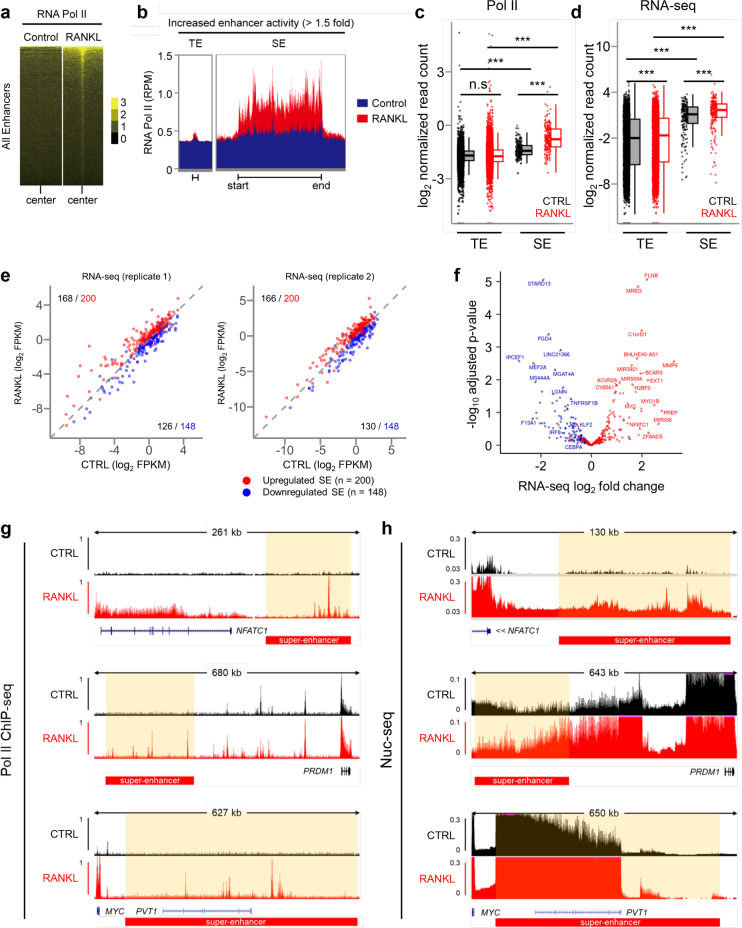
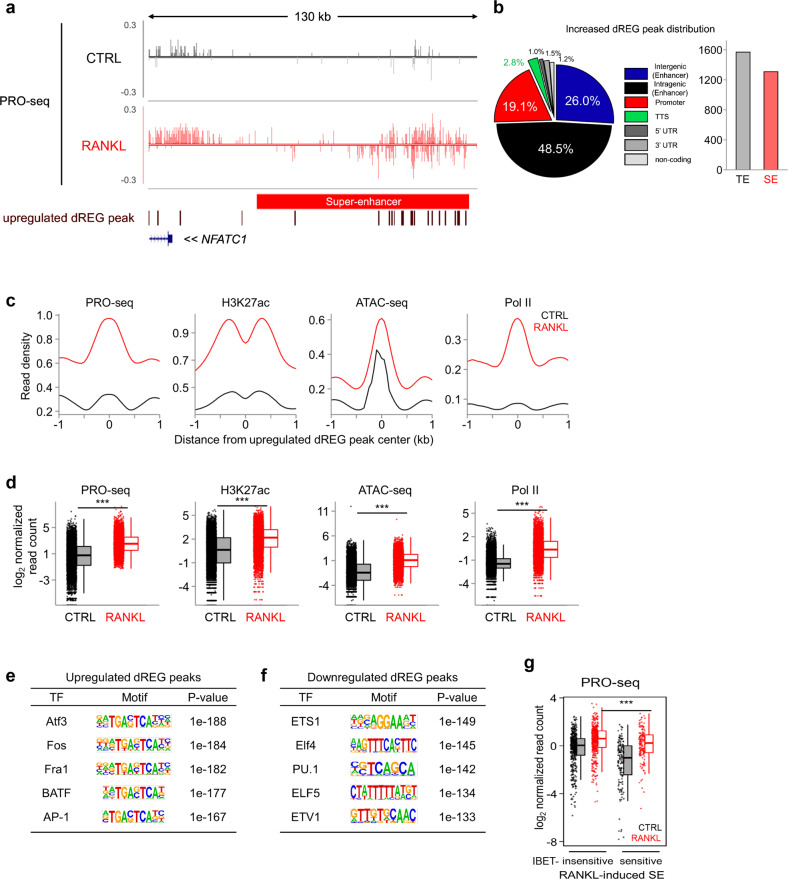
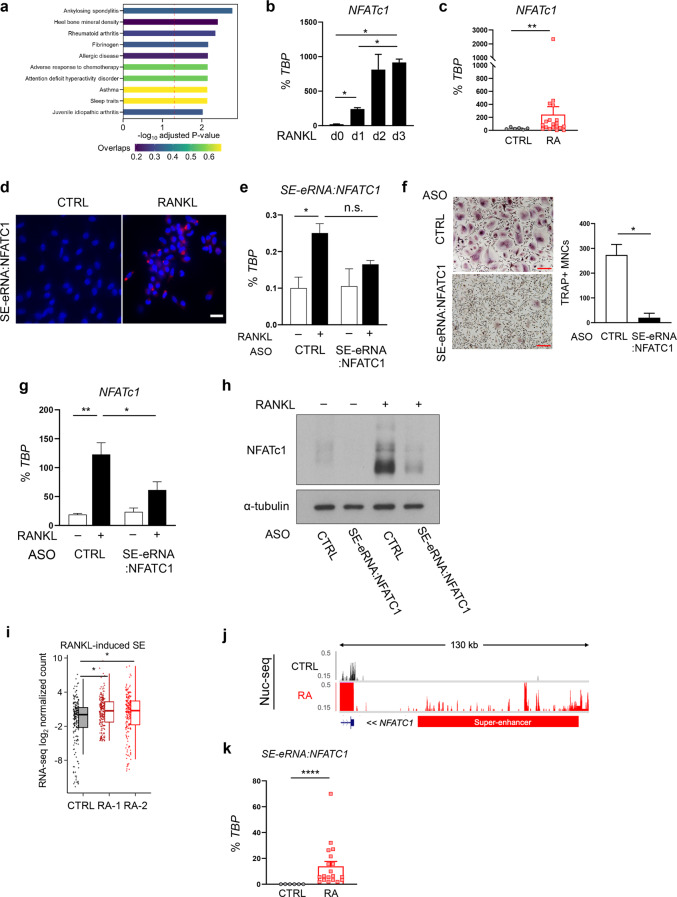
Monocyte/macrophage lineage cells are highly plastic and can differentiate into various cells under different environmental stimuli. Bone-resorbing osteoclasts are derived from the monocyte/macrophage lineage in response to receptor activator of NF-κB ligand (RANKL). However, the epigenetic signature contributing to the fate commitment of monocyte/macrophage lineage differentiation into human osteoclasts is largely unknown. In this study, we identified RANKL-responsive human osteoclast-specific superenhancers (SEs) and SE-associated enhancer RNAs (SE-eRNAs) by integrating data obtained from ChIP-seq, ATAC-seq, nuclear RNA-seq and PRO-seq analyses. RANKL induced the formation of 200 SEs, which are large clusters of enhancers, while suppressing 148 SEs in macrophages. RANKL-responsive SEs were strongly correlated with genes in the osteoclastogenic program and were selectively increased in human osteoclasts but marginally presented in osteoblasts, CD4+ T cells, and CD34+ cells. In addition to the major transcription factors identified in osteoclasts, we found that BATF binding motifs were highly enriched in RANKL-responsive SEs. The depletion of BATF1/3 inhibited RANKL-induced osteoclast differentiation. Furthermore, we found increased chromatin accessibility in SE regions, where RNA polymerase II was significantly recruited to induce the extragenic transcription of SE-eRNAs, in human osteoclasts. Knocking down SE-eRNAs in the vicinity of the NFATc1 gene diminished the expression of NFATc1, a major regulator of osteoclasts, and osteoclast differentiation. Inhibiting BET proteins suppressed the formation of some RANKL-responsive SEs and NFATc1-associated SEs, and the expression of SE-eRNA:NFATc1. Moreover, SE-eRNA:NFATc1 was highly expressed in the synovial macrophages of rheumatoid arthritis patients exhibiting high-osteoclastogenic potential. Our genome-wide analysis revealed RANKL-inducible SEs and SE-eRNAs as osteoclast-specific signatures, which may contribute to the development of osteoclast-specific therapeutic interventions.
Keywords: Osteoclasts; Rheumatoid arthritis; enhancer RNAs; super-enhancers.
© 2022. The Author(s), under exclusive licence to CSI and USTC.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
