

Progressive differentiation toward the long-lived plasma cell compartment in the bone marrow
- PMID: 36515679
- PMCID: PMC9754767
- DOI: 10.1084/jem.20221717
Progressive differentiation toward the long-lived plasma cell compartment in the bone marrow
Abstract
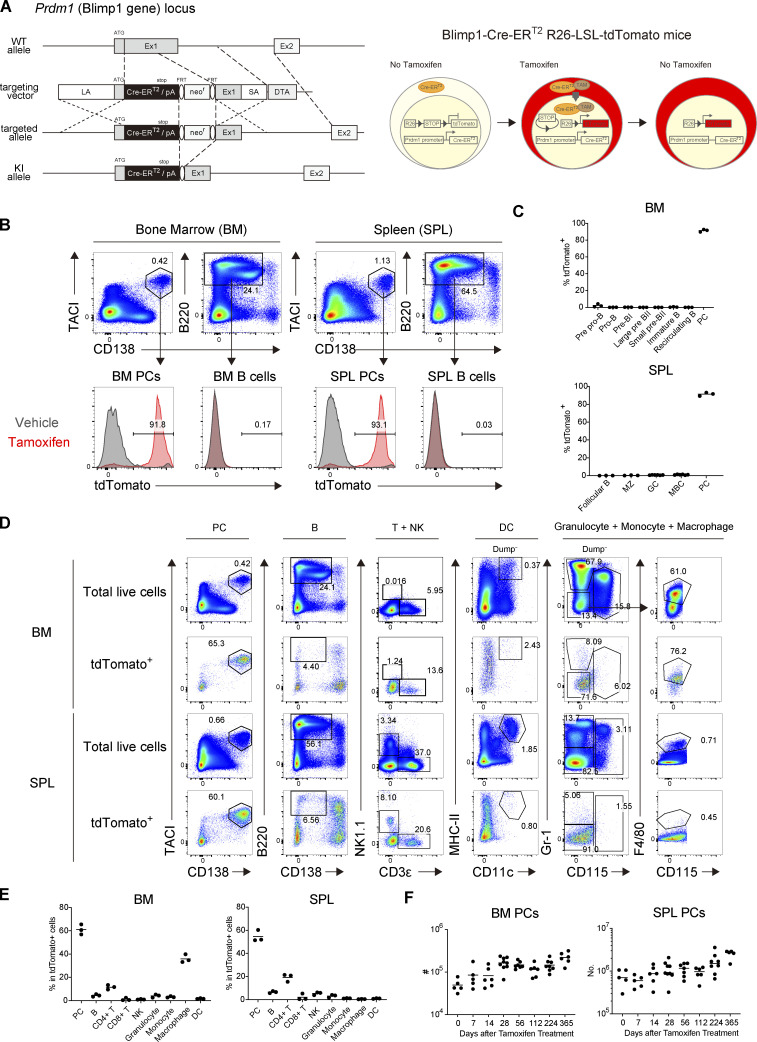
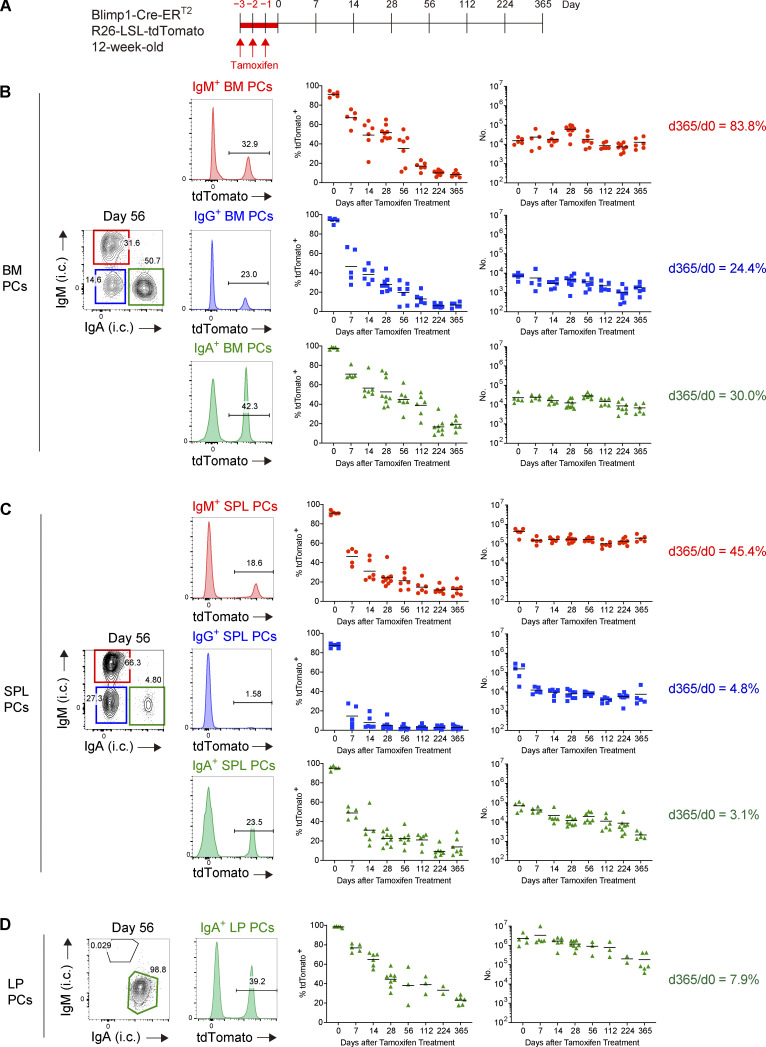
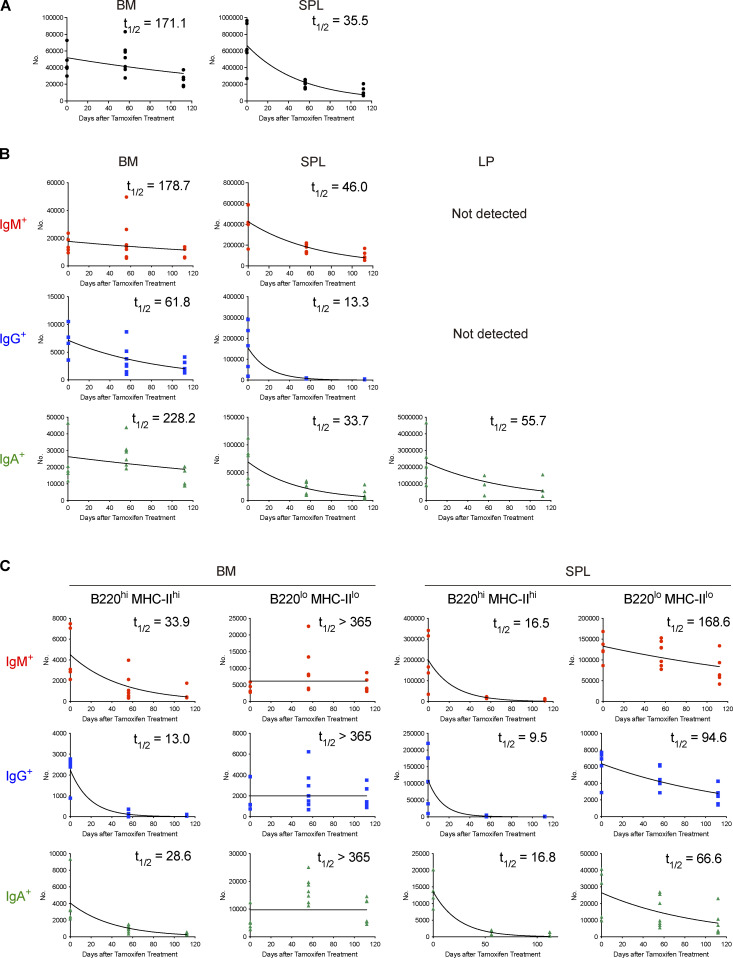
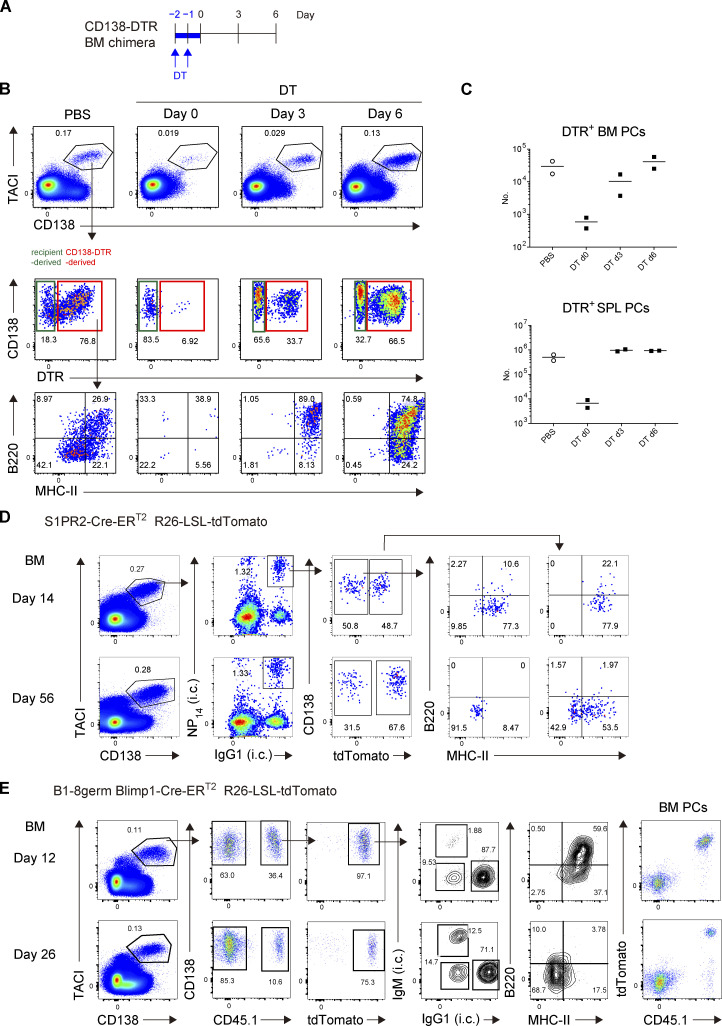
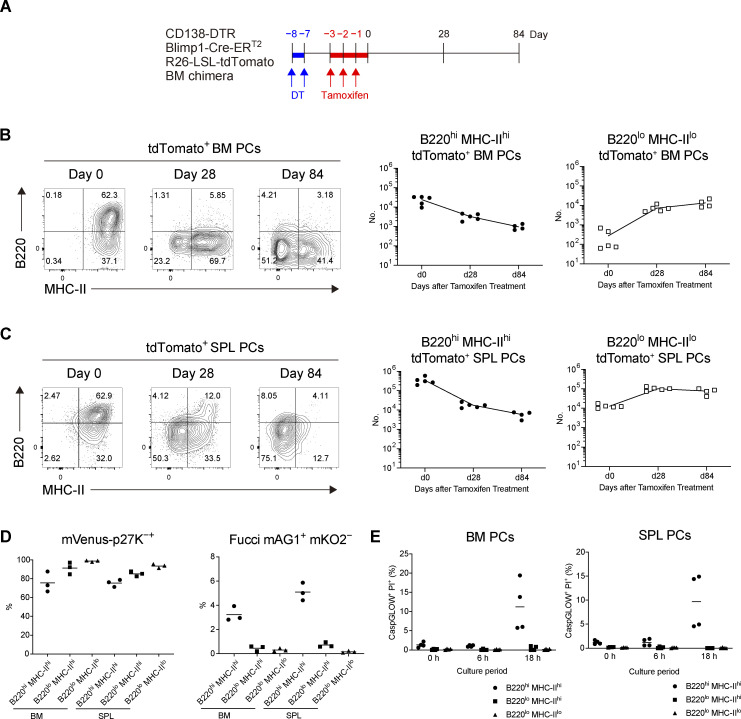
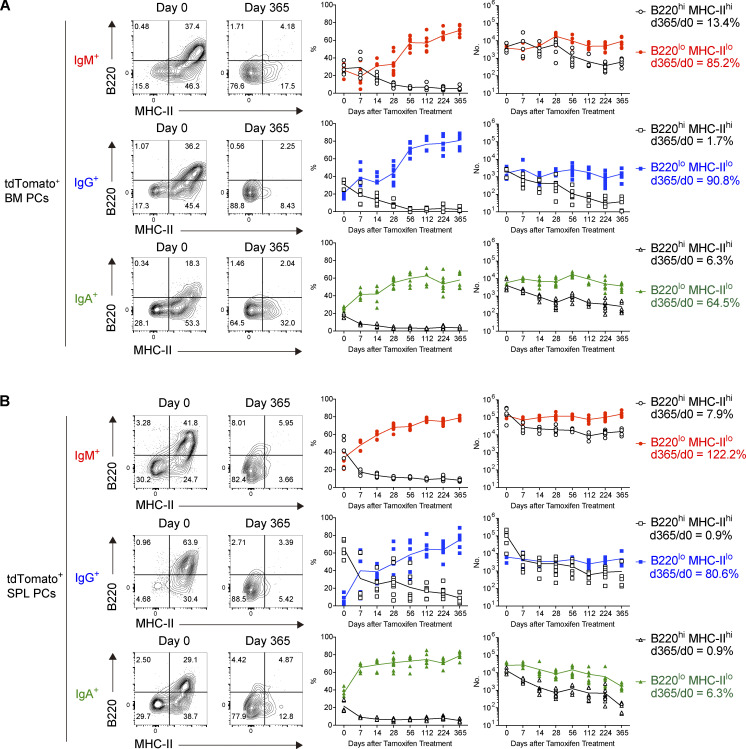
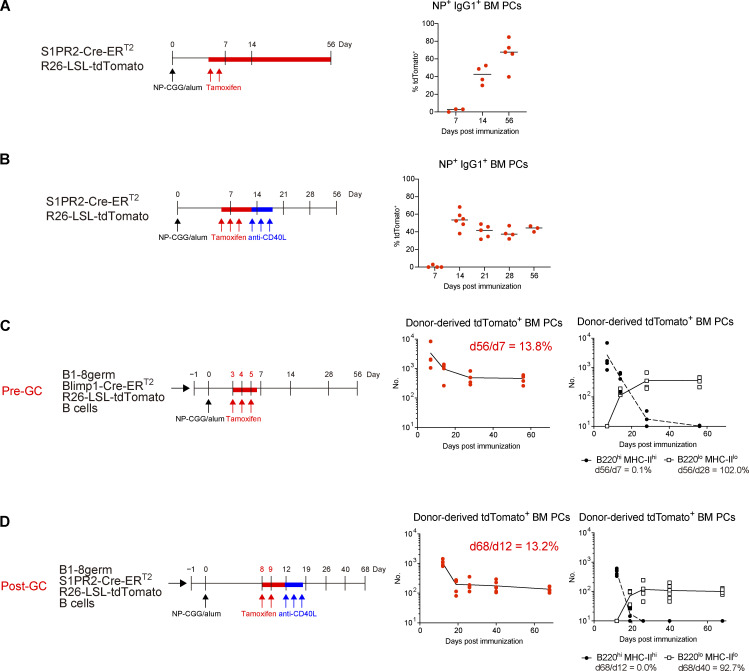
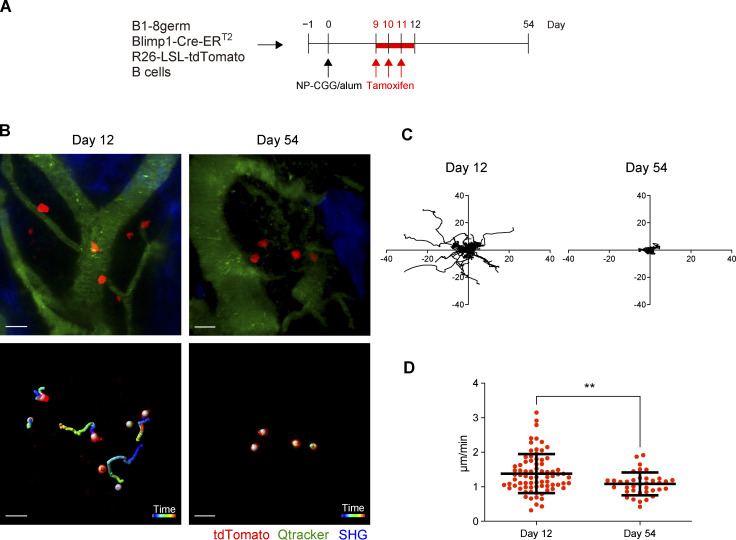
The longevity of plasma cells is dependent on their ability to access and reside in so-called niches that are predominantly located in the bone marrow. Here, by employing a traceable method to label recently generated plasma cells, we showed that homeostatic plasma cells in the bone marrow and spleen were continuously replenished by newly generated B220hiMHC-IIhi populations that progressively differentiated into B220loMHC-IIlo long-lived plasma cell (LLPC) populations. We also found that, in the bone marrow, germinal center (GC)-independent and GC-dependent plasma cells decayed similarly upon NP-CGG engagement, and both entered the B220loMHC-IIlo LLPC pool. Compared with NP+B220hiMHC-IIhi plasma cells, NP+B220loMHC-IIlo cells were more immobilized in the bone marrow niches and showed better survival potential. Thus, our results suggest that the adhesion status of bone marrow plasma cells is dynamically altered during their differentiation and is associated with provision of survival signals.
© 2022 Koike et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
