

Fibroblast Activation Protein Activates Macrophages and Promotes Parenchymal Liver Inflammation and Fibrosis
- PMID: 36521660
- PMCID: PMC9972574
- DOI: 10.1016/j.jcmgh.2022.12.005
Fibroblast Activation Protein Activates Macrophages and Promotes Parenchymal Liver Inflammation and Fibrosis
Abstract
Background & aims: Fibroblast activation protein (FAP) is expressed on activated fibroblast. Its role in fibrosis and desmoplasia is controversial, and data on pharmacological FAP inhibition are lacking. We aimed to better define the role of FAP in liver fibrosis in vivo and in vitro.
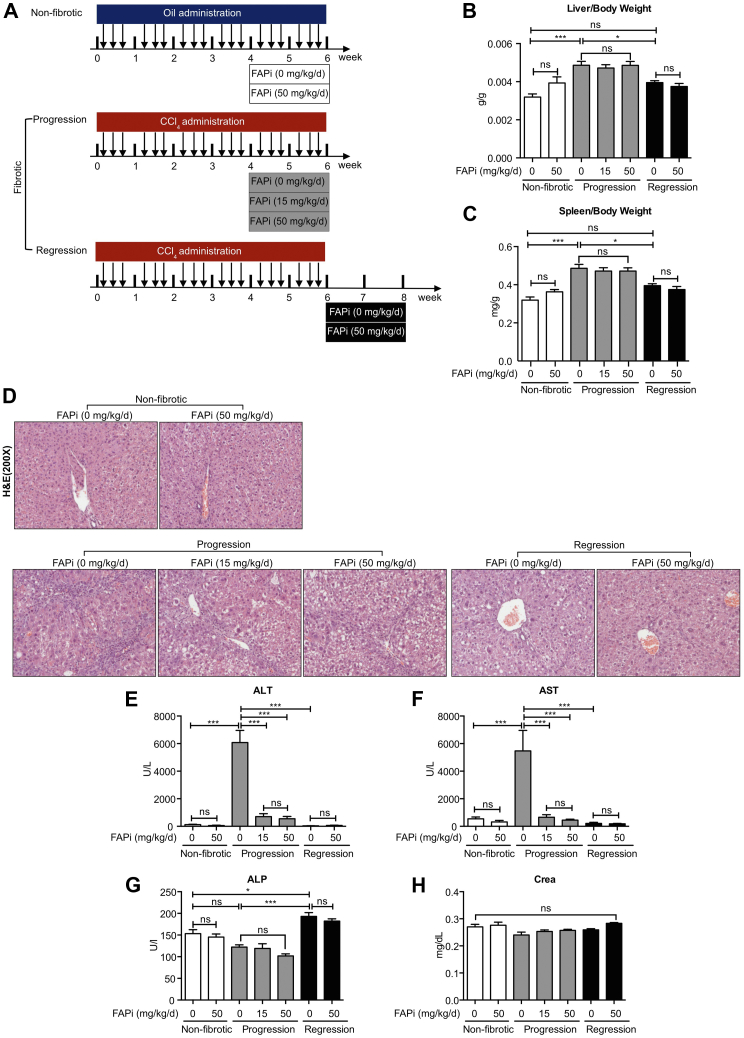
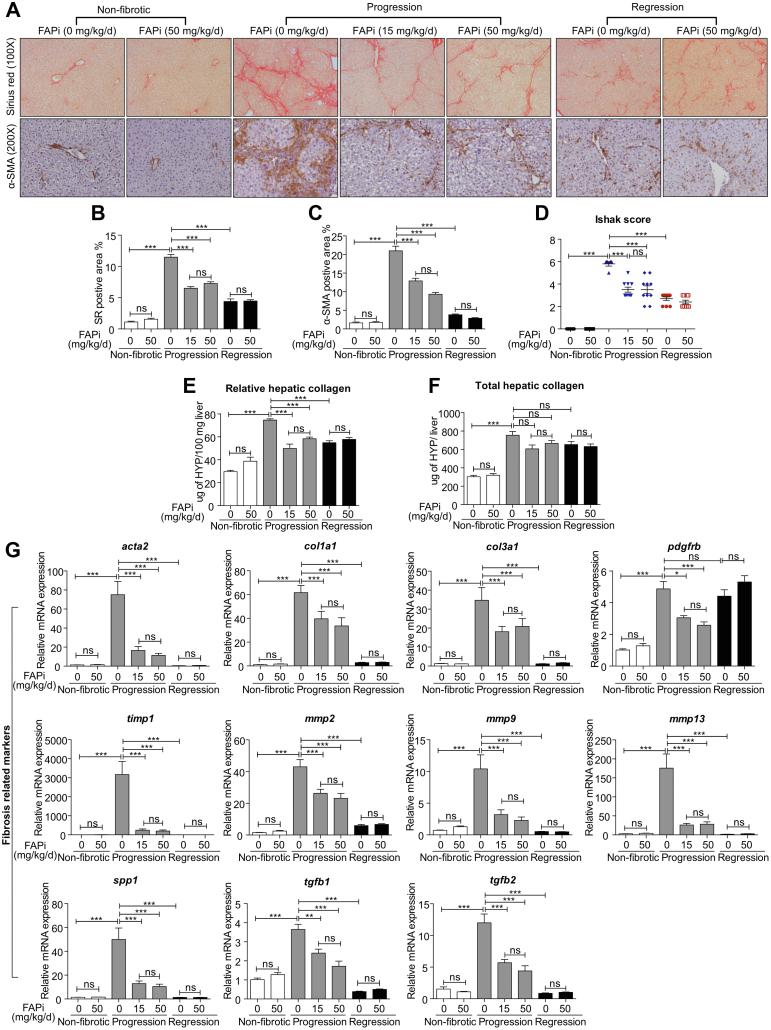
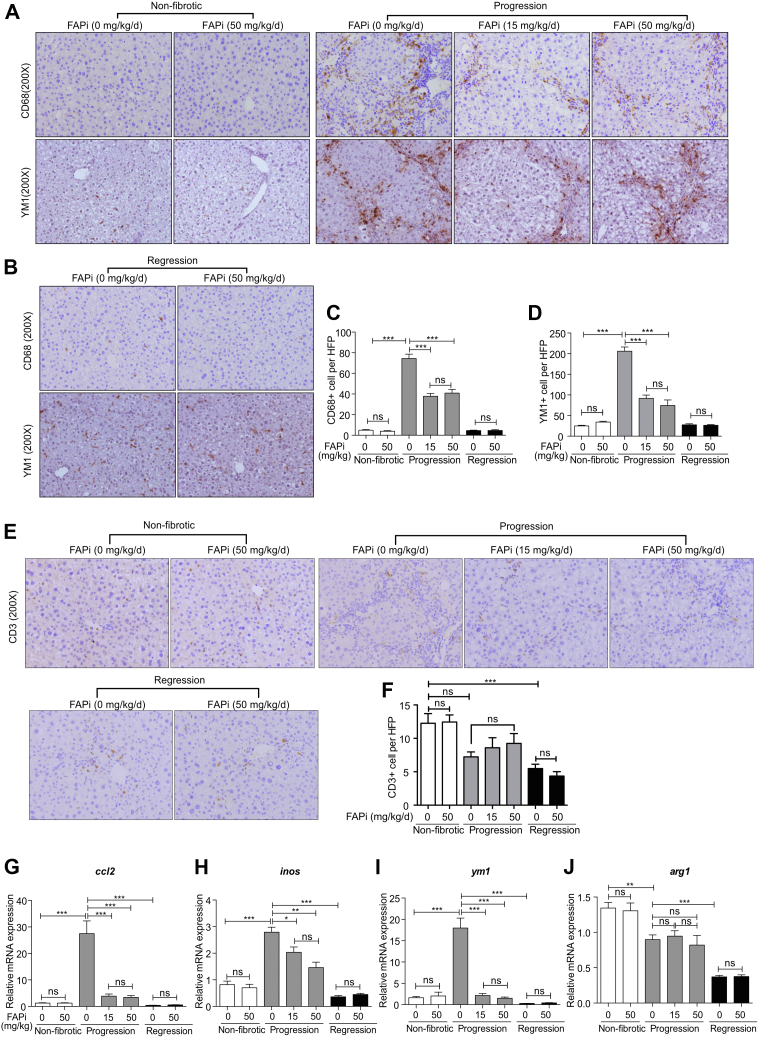
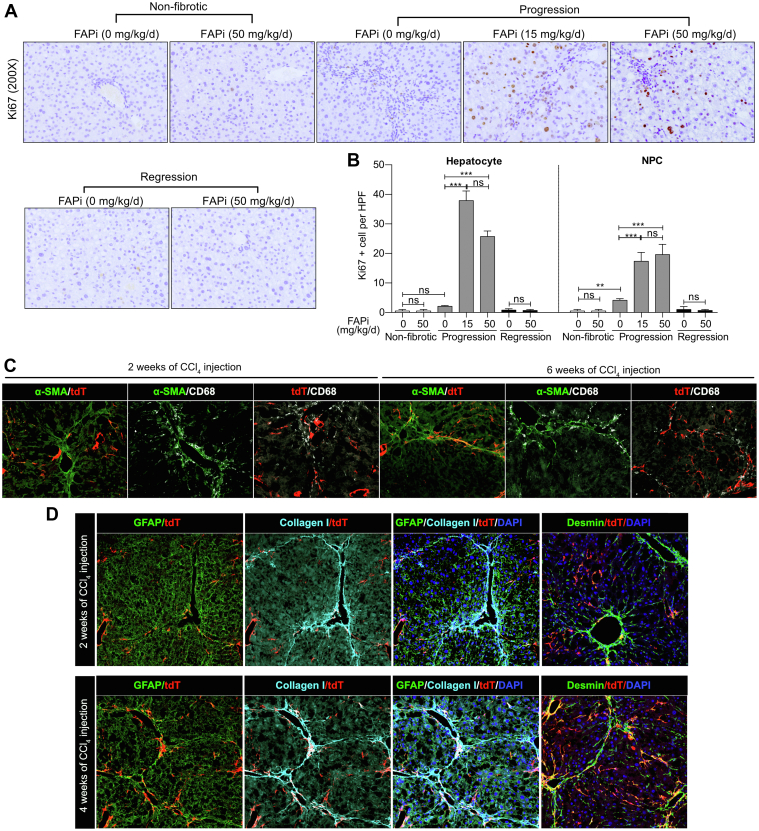
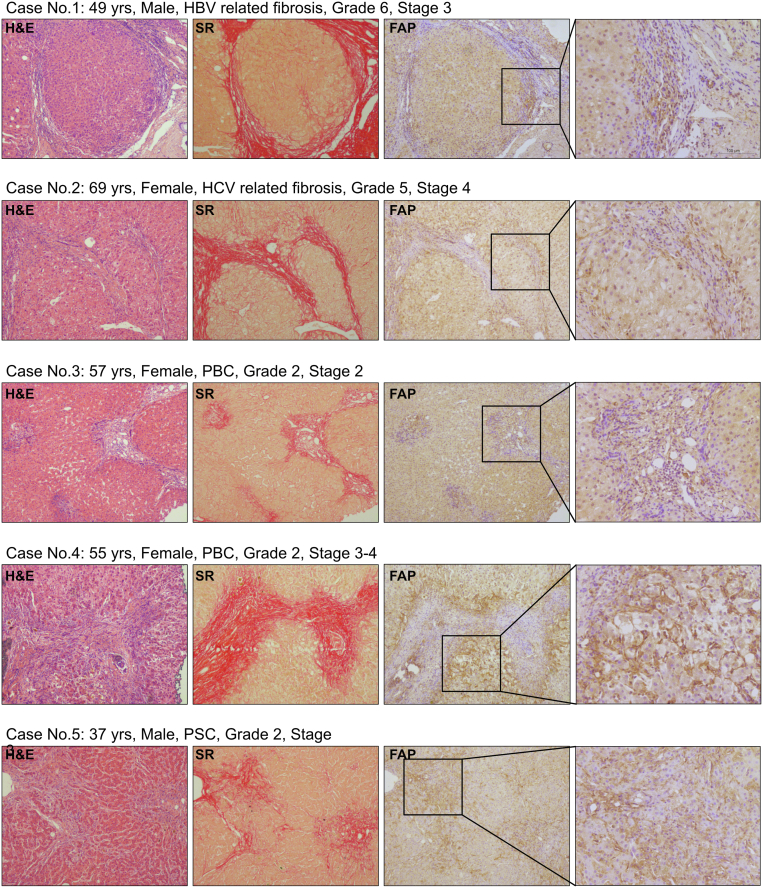
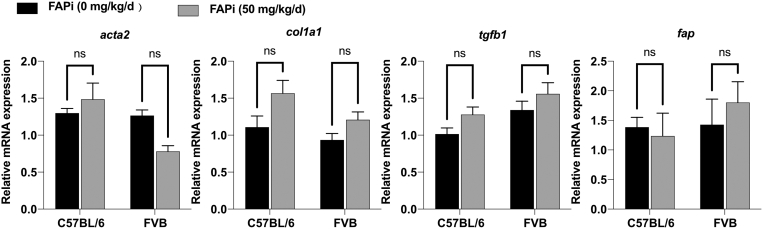
Methods: FAP expression was analyzed in mice and patients with fibrotic liver diseases of various etiologies. Fibrotic mice received a specific FAP inhibitor (FAPi) at 2 doses orally for 2 weeks during parenchymal fibrosis progression (6 weeks of carbon tetrachloride) and regression (2 weeks off carbon tetrachloride), and with biliary fibrosis (Mdr2-/-). Recombinant FAP was added to (co-)cultures of hepatic stellate cells (HSC), fibroblasts, and macrophages. Fibrosis- and inflammation-related parameters were determined biochemically, by quantitative immunohistochemistry, polymerase chain reaction, and transcriptomics.
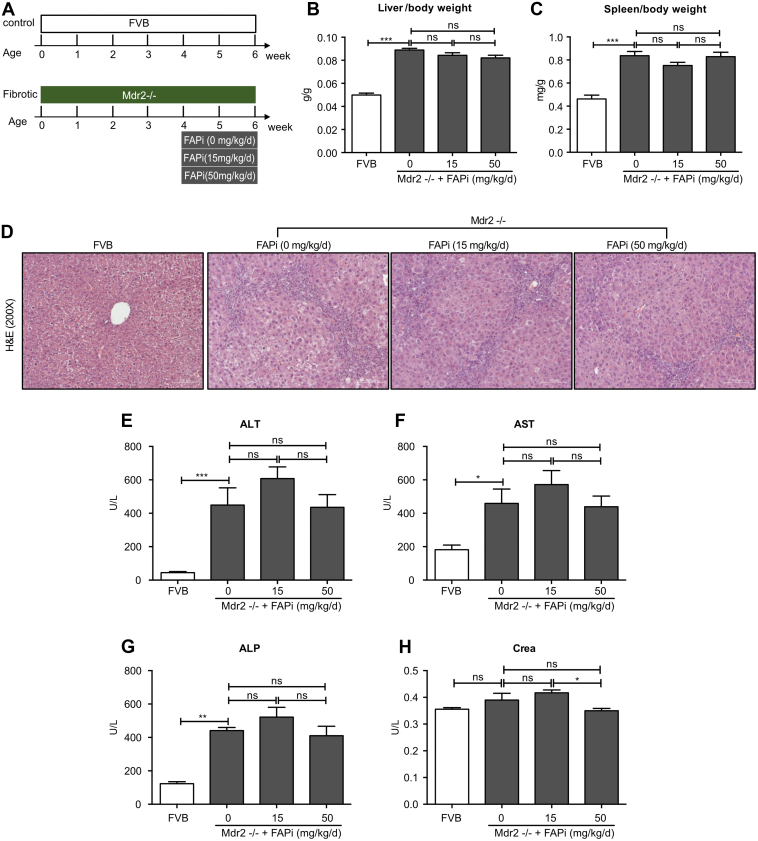
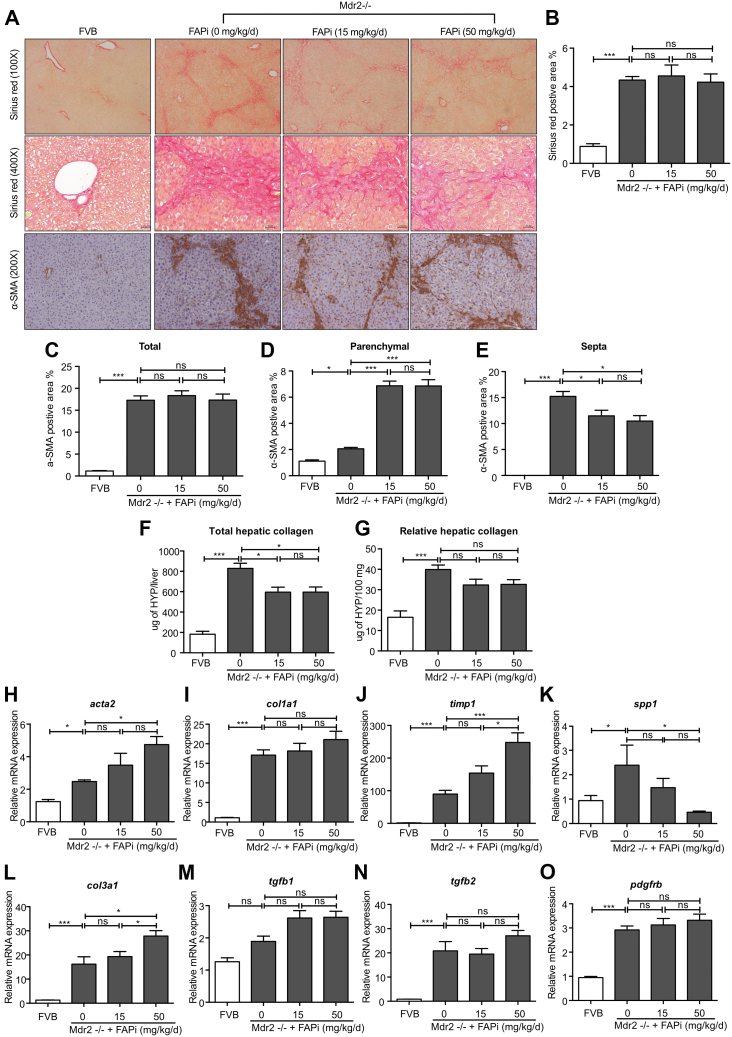
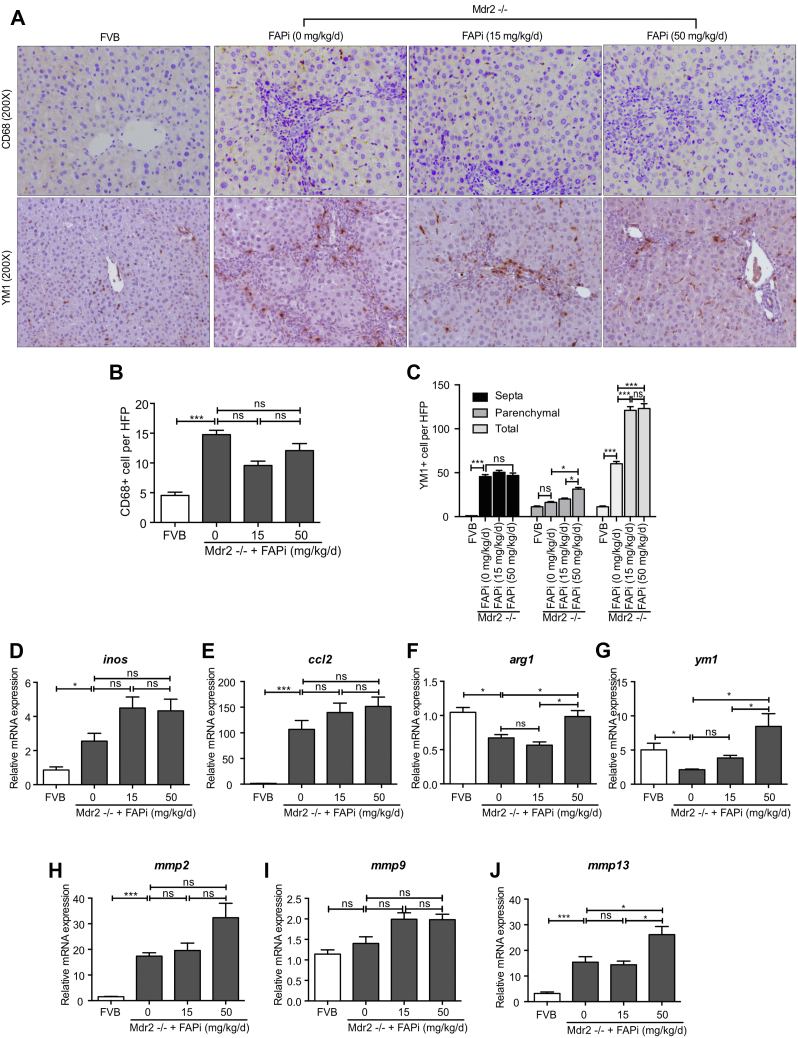
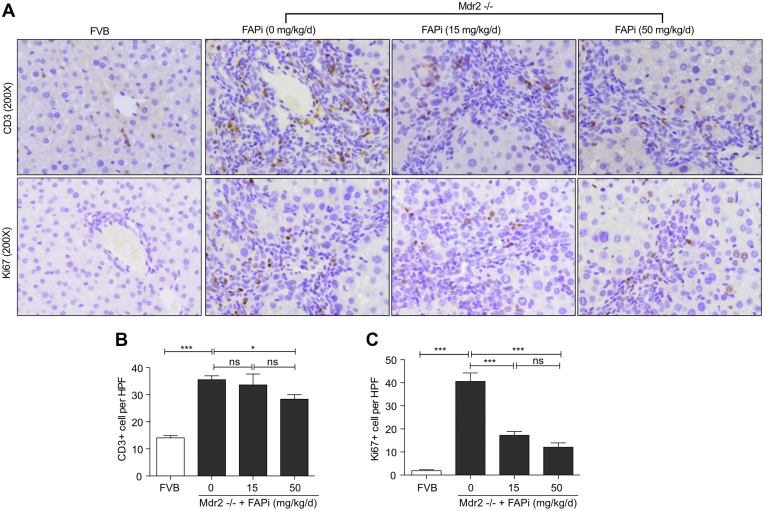
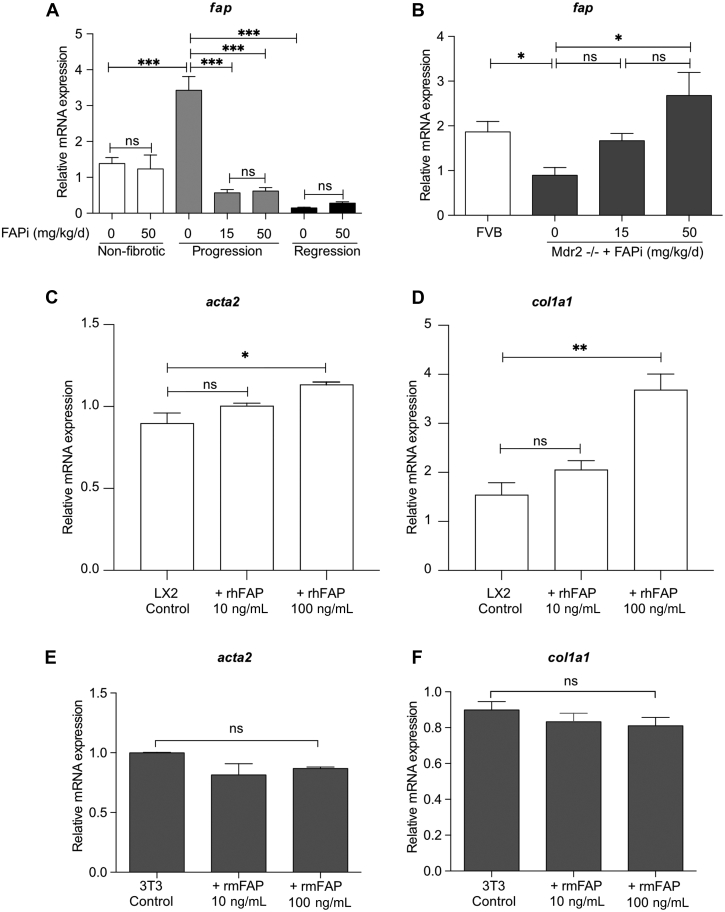
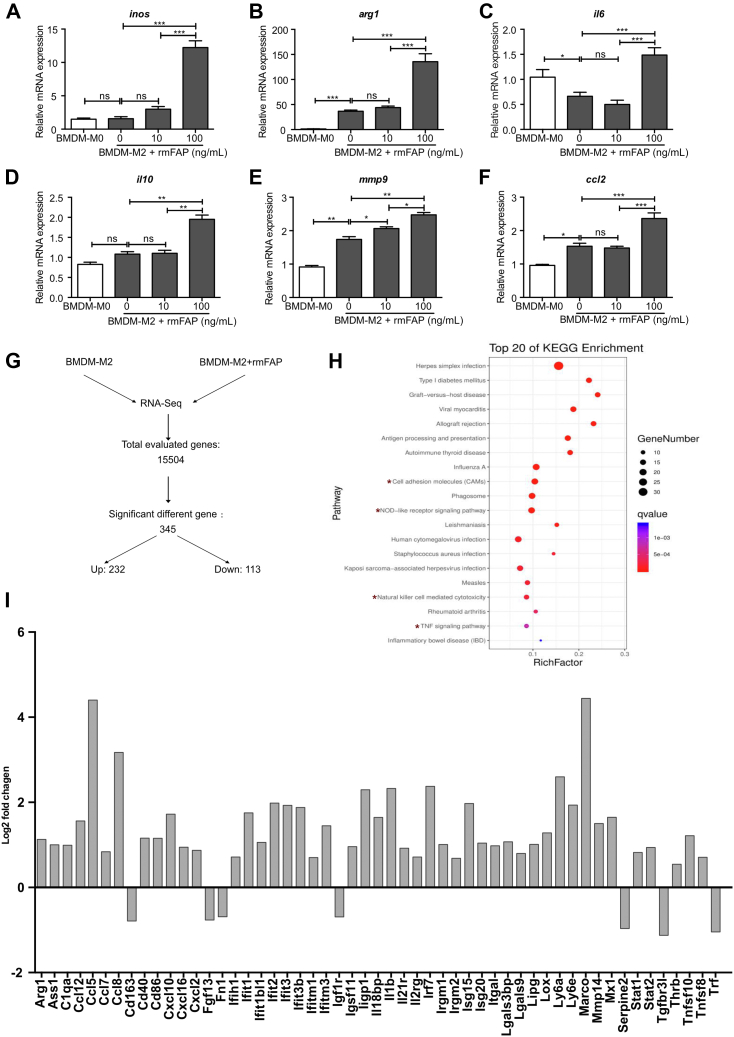
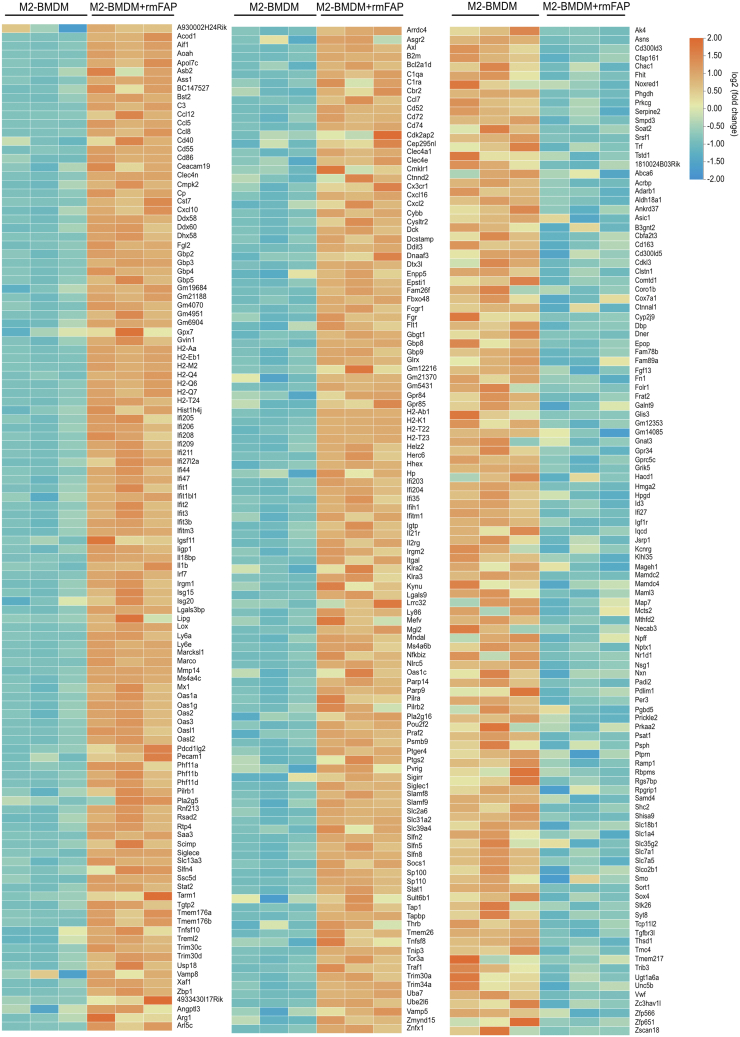
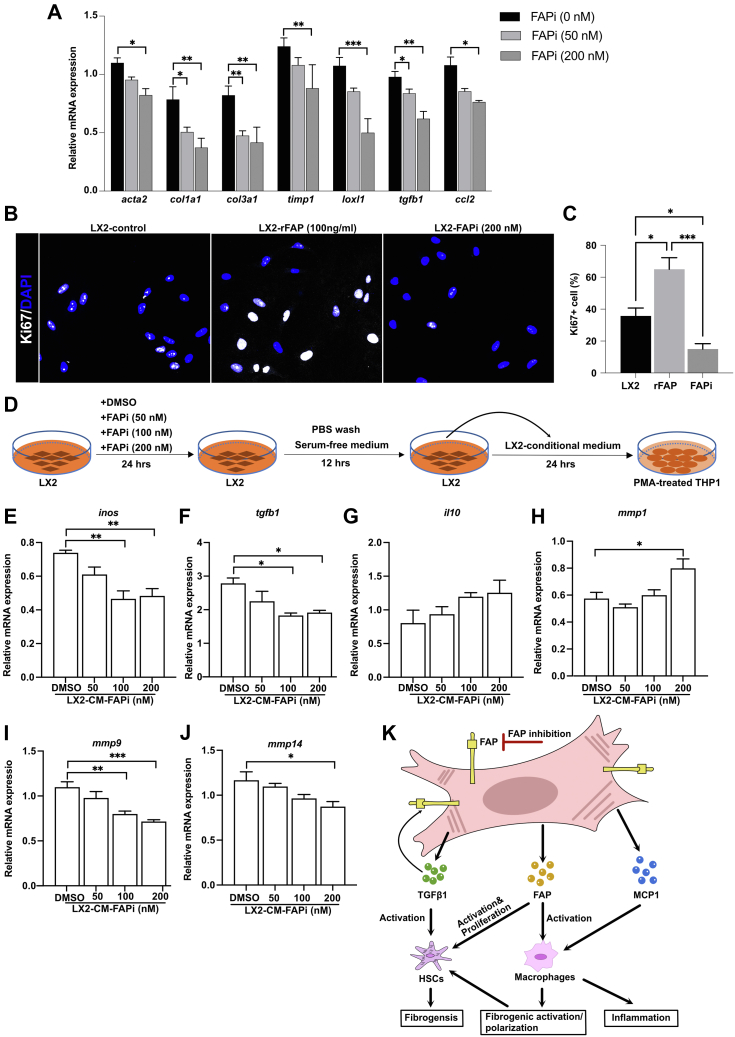
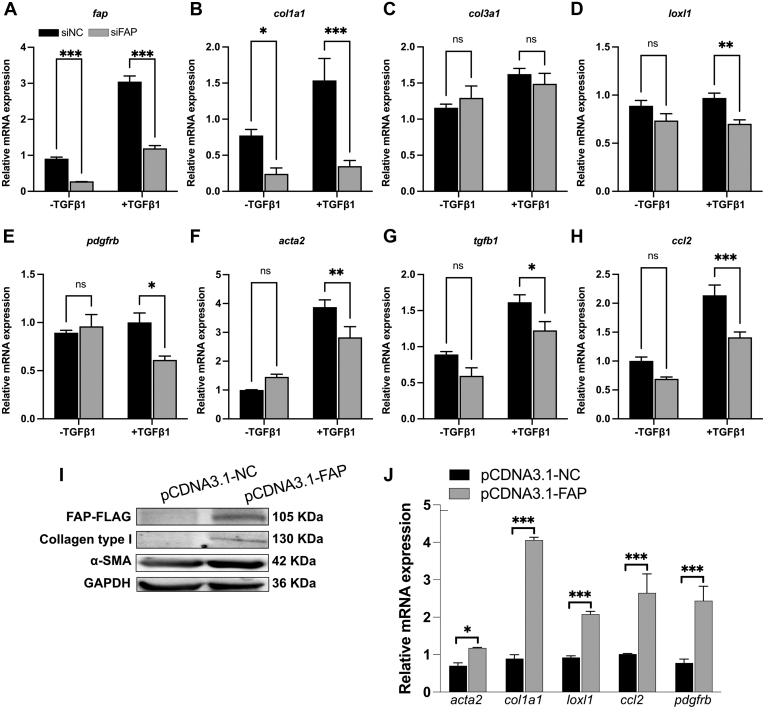
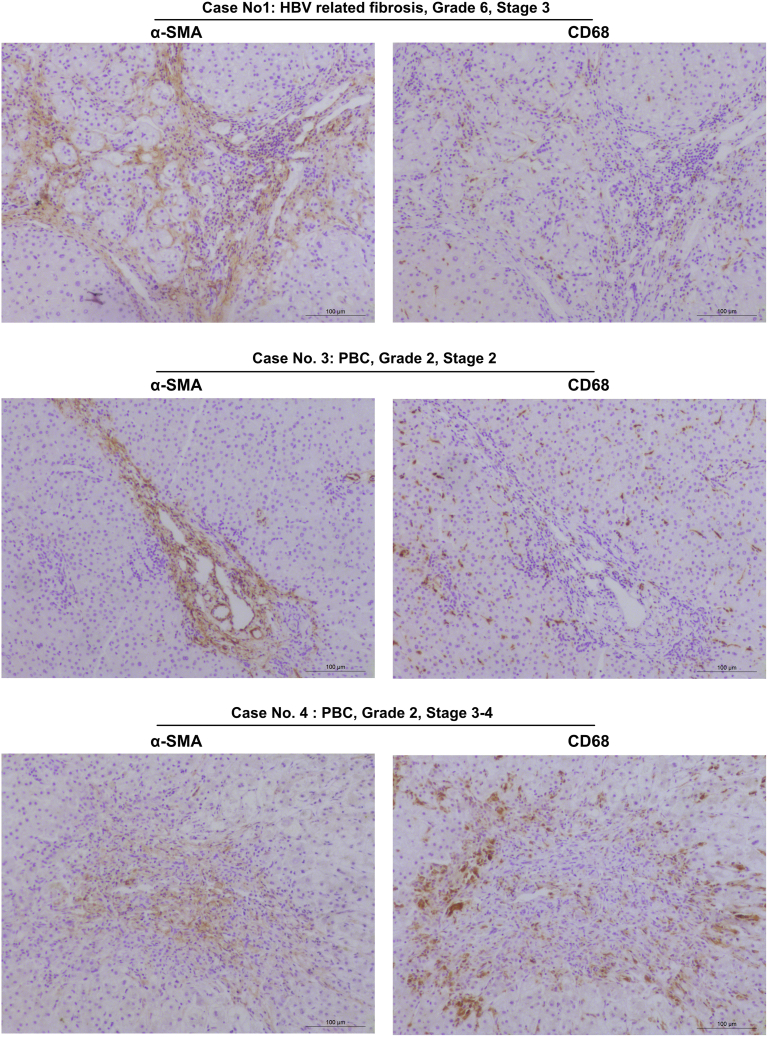
Results: FAP+ fibroblasts/HSCs were α-smooth muscle actin (α-SMA)-negative and located at interfaces of fibrotic septa next to macrophages in murine and human livers. In parenchymal fibrosis, FAPi reduced collagen area, liver collagen content, α-SMA+ myofibroblasts, M2-type macrophages, serum alanine transaminase and aspartate aminotransferase, key fibrogenesis-related transcripts, and increased hepatocyte proliferation 10-fold. During regression, FAP was suppressed, and FAPi was ineffective. FAPi less potently inhibited biliary fibrosis. In vitro, FAP small interfering RNA reduced HSC α-SMA expression and collagen production, and FAPi suppressed their activation and proliferation. Compared with untreated macrophages, FAPi regulated macrophage profibrogenic activation and transcriptome, and their conditioned medium attenuated HSC activation, which was increased with addition of recombinant FAP.
Conclusions: Pharmacological FAP inhibition attenuates inflammation-predominant liver fibrosis. FAP is expressed on subsets of activated fibroblasts/HSC and promotes both macrophage and HSC profibrogenic activity in liver fibrosis.
Keywords: Antifibrotic Therapy; Fibroblast Activation Protein (FAP); Hepatic Stellate Cell; Liver Fibrosis; Macrophage.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
FAP: Not Just a Biomarker but Druggable Target in Liver Fibrosis.Cell Mol Gastroenterol Hepatol. 2023;15(4):1018-1019. doi: 10.1016/j.jcmgh.2022.12.018. Epub 2023 Jan 18. Cell Mol Gastroenterol Hepatol. 2023. PMID: 36681094 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
