

Epigenetic regulation of IPF fibroblast phenotype by glutaminolysis
- PMID: 36526153
- PMCID: PMC9827063
- DOI: 10.1016/j.molmet.2022.101655
Epigenetic regulation of IPF fibroblast phenotype by glutaminolysis
Abstract
Objective: Excessive extra-cellular-matrix production and uncontrolled proliferation of the fibroblasts are characteristics of many fibrotic diseases, including idiopathic pulmonary fibrosis (IPF). The fibroblasts have enhanced glutaminolysis with up-regulated glutaminase, GLS1, which converts glutamine to glutamate. Here, we investigated the role of glutaminolysis and glutaminolysis-derived metabolite α-ketoglutarate (α-KG) on IPF fibroblast phenotype and gene expression.
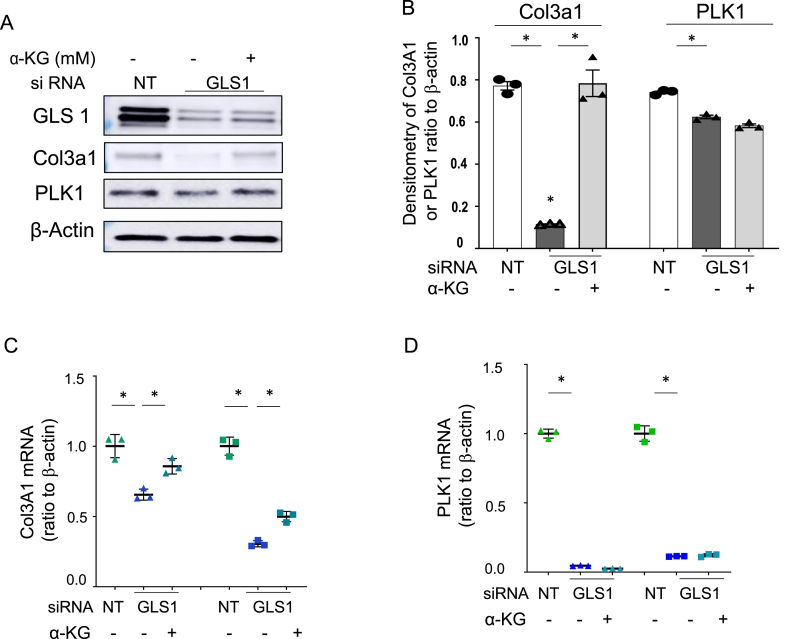
Methods: Reduced glutamine conditions were carried out either using glutamine-free culture medium or silencing the expression of GLS1 with siRNA, with or without α-KG compensation. Cell phenotype has been characterized under these different conditions, and gene expression profile was examined by RNA-Seq. Specific profibrotic genes (Col3A1 and PLK1) expression were examined by real-time PCR and western blots. The levels of repressive histone H3K27me3, which demethylase activity is affected by glutaminolysis, were examined and H3K27me3 association with promoter region of Col3A1 and PLK1 were checked by ChIP assays. Effects of reduced glutaminolysis on fibrosis markers were checked in an animal model of lung fibrosis.
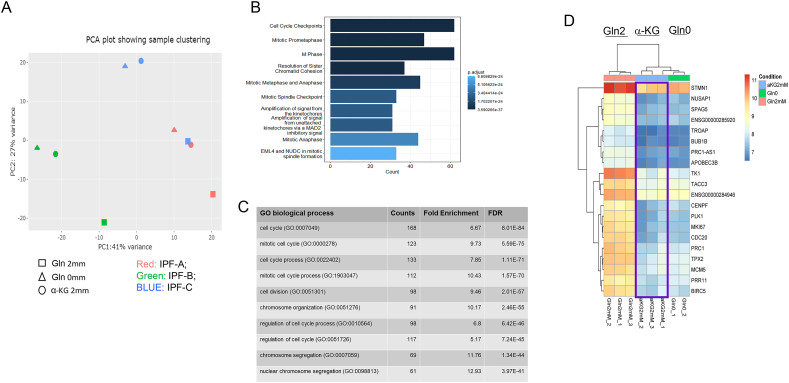
Results: The lack of glutamine in the culture medium alters the profibrotic phenotype of activated fibroblasts. The addition of exogenous and glutaminolysis-derived metabolite α-KG to glutamine-free media barely restores the pro-fibrotic phenotype of activated fibroblasts. Many genes are down-regulated in glutamine-free medium, α-KG supplementation only rescues a limited number of genes. As α-KG is a cofactor for histone demethylases of H3K27me3, the reduced glutaminolysis alters H3K27me3 levels, and enriches H3K27me3 association with Col3A1 and PLK1 promoter region. Adding α-KG in glutamine-free medium depleted H3K27me3 association with Col3A1 promoter region but not that of PLK1. In a murine model of lung fibrosis, mice with reduced glutaminolysis showed markedly reduced fibrotic markers.
Conclusions: This study indicates that glutamine is critical for supporting pro-fibrotic fibroblast phenotype in lung fibrosis, partially through α-KG-dependent and -independent mechanisms, and supports targeting fibroblast metabolism as a therapeutic method for fibrotic diseases.
Keywords: Col3A1; Glutamine; H3K27me3; Lung fibrosis; PLK1; Running head; α-KG.
Copyright © 2022 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Figures




References
-
- Smith R.J. Glutamine metabolism and its physiologic importance. JPEN J Parenter Enteral Nutr. 1990;14(4 Suppl):40S–44S. - PubMed
-
- Neu J, Demarco V, Freeman B, McCain M, Strauss D. Is glutamine a conditionally essential amino acid during pregnancy?177. Pediatr Res. 1996;40:544.
-
- Jackson NC, Carroll PV, Russell-Jones DL, Sonksen PH, Treacher DF, Umpleby AM. The metabolic consequences of critical illness: acute effects on glutamine and protein metabolism. Am J Physiol. 1999;276(1 Pt 1):E163–E170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
