

A mean-field model of glutamate and GABA synaptic dynamics for functional MRS
- PMID: 36528313
- PMCID: PMC7614487
- DOI: 10.1016/j.neuroimage.2022.119813
A mean-field model of glutamate and GABA synaptic dynamics for functional MRS
Abstract
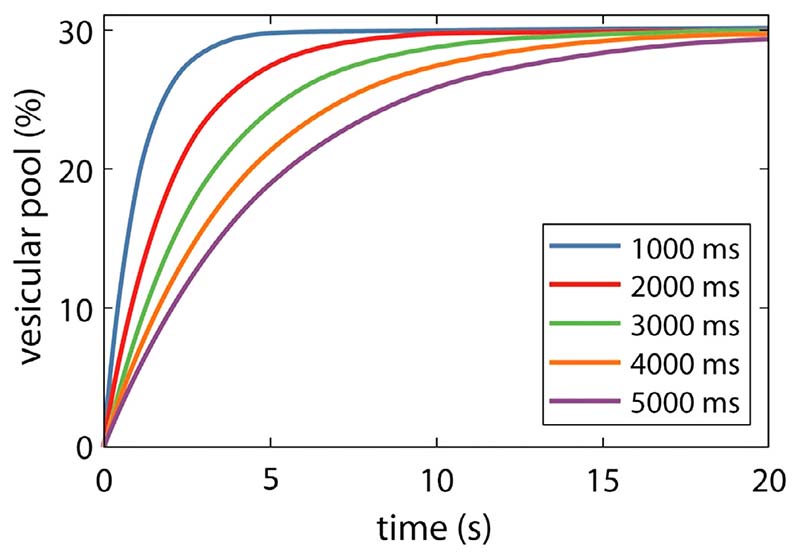
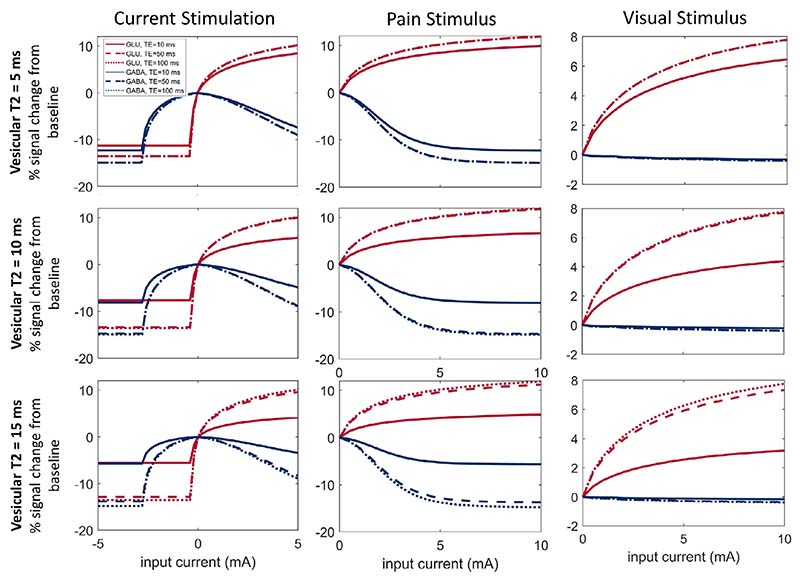
Advances in functional magnetic resonance spectroscopy (fMRS) have enabled the quantification of activity-dependent changes in neurotransmitter concentrations in vivo. However, the physiological basis of the large changes in GABA and glutamate observed by fMRS (>10%) over short time scales of less than a minute remain unclear as such changes cannot be accounted for by known synthesis or degradation metabolic pathways. Instead, it has been hypothesized that fMRS detects shifts in neurotransmitter concentrations as they cycle from presynaptic vesicles, where they are largely invisible, to extracellular and cytosolic pools, where they are detectable. The present paper uses a computational modelling approach to demonstrate the viability of this hypothesis. A new mean-field model of the neural mechanisms generating the fMRS signal in a cortical voxel is derived. The proposed macroscopic mean-field model is based on a microscopic description of the neurotransmitter dynamics at the level of the synapse. Specifically, GABA and glutamate are assumed to cycle between three metabolic pools: packaged in the vesicles; active in the synaptic cleft; and undergoing recycling and repackaging in the astrocytic or neuronal cytosol. Computational simulations from the model are used to generate predicted changes in GABA and glutamate concentrations in response to different types of stimuli including pain, vision, and electric current stimulation. The predicted changes in the extracellular and cytosolic pools corresponded to those reported in empirical fMRS data. Furthermore, the model predicts a selective control mechanism of the GABA/glutamate relationship, whereby inhibitory stimulation reduces both neurotransmitters, whereas excitatory stimulation increases glutamate and decreases GABA. The proposed model bridges between neural dynamics and fMRS and provides a mechanistic account for the activity-dependent changes in the glutamate and GABA fMRS signals. Lastly, these results indicate that echo-time may be an important timing parameter that can be leveraged to maximise fMRS experimental outcomes.
Keywords: FMRS; GABA; Glutamate; Magnetic resonance spectroscopy; Mathematical modelling; Mean field model.
Copyright © 2022. Published by Elsevier Inc.
Figures






References
-
- An H, Lin W. Spin Density, T1, T2, T2 ∗ Relaxation and Bloch Equations. Current Protocols in Magnetic Resonance Imaging. 2001;00:B3.1.1–B3.1.10. doi: 10.1002/0471142719.mib0301s00. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
