

Saturated very long-chain fatty acids regulate macrophage plasticity and invasiveness
- PMID: 36528616
- PMCID: PMC9759912
- DOI: 10.1186/s12974-022-02664-y
Saturated very long-chain fatty acids regulate macrophage plasticity and invasiveness
Abstract
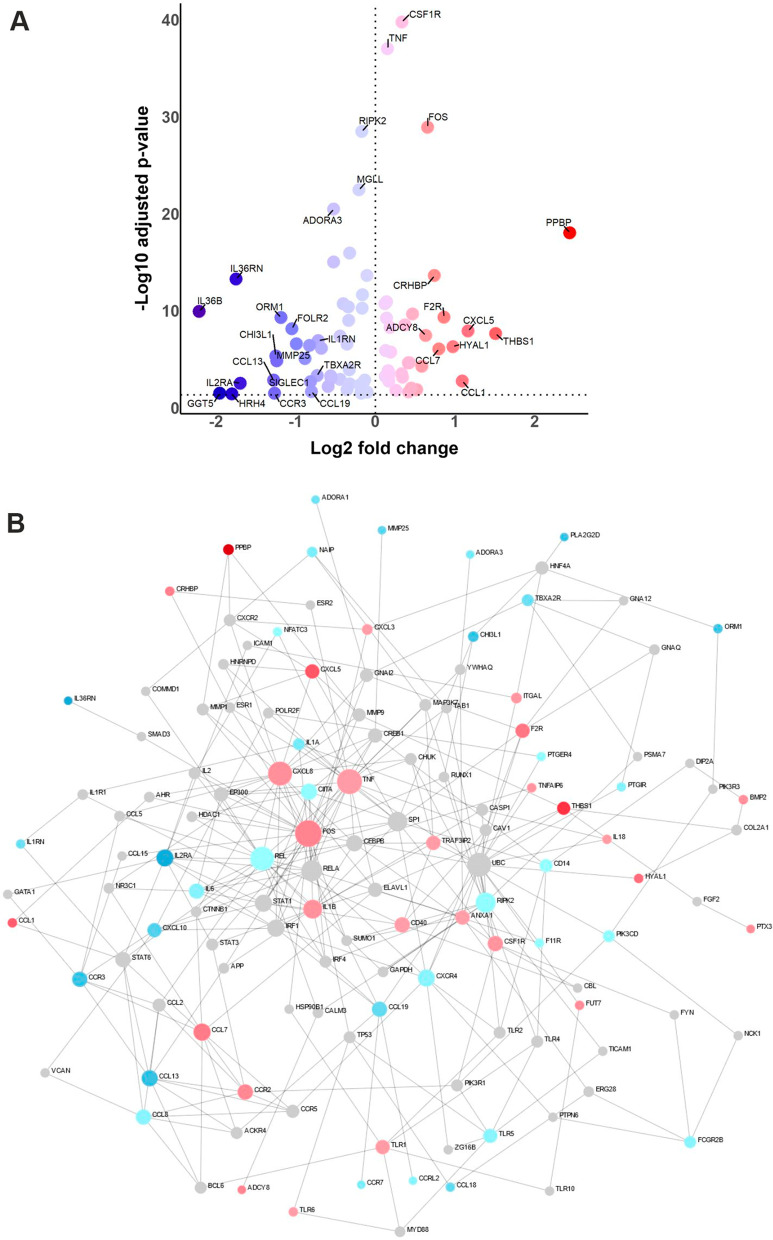
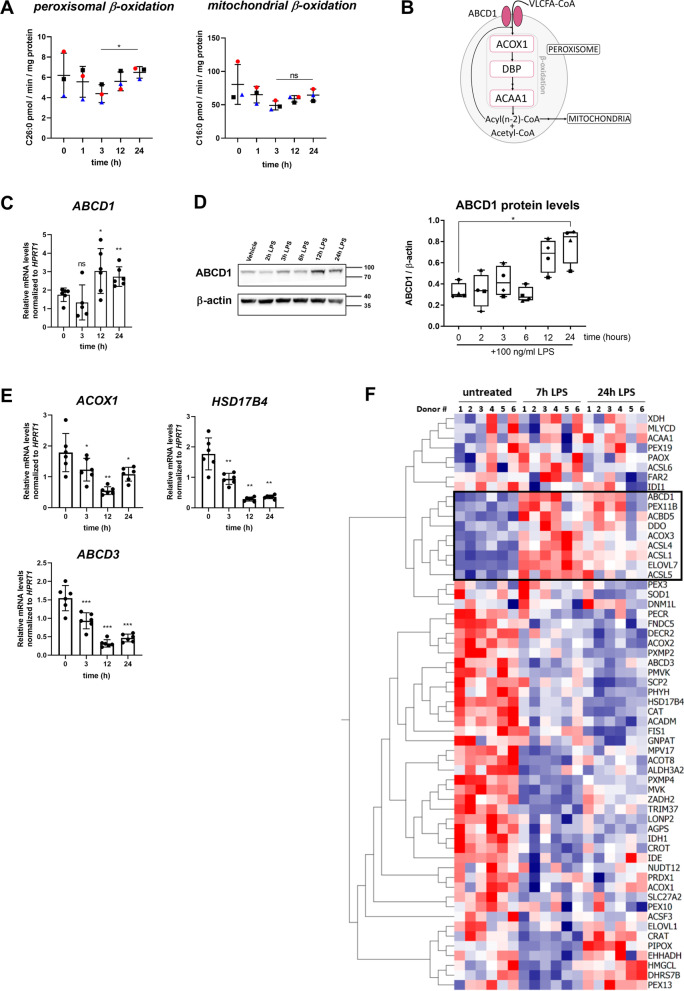
Saturated very long-chain fatty acids (VLCFA, ≥ C22), enriched in brain myelin and innate immune cells, accumulate in X-linked adrenoleukodystrophy (X-ALD) due to inherited dysfunction of the peroxisomal VLCFA transporter ABCD1. In its severest form, X-ALD causes cerebral myelin destruction with infiltration of pro-inflammatory skewed monocytes/macrophages. How VLCFA levels relate to macrophage activation is unclear. Here, whole transcriptome sequencing of X-ALD macrophages indicated that VLCFAs prime human macrophage membranes for inflammation and increased expression of factors involved in chemotaxis and invasion. When added externally to mimic lipid release in demyelinating X-ALD lesions, VLCFAs did not activate toll-like receptors in primary macrophages. In contrast, VLCFAs provoked pro-inflammatory responses through scavenger receptor CD36-mediated uptake, cumulating in JNK signalling and expression of matrix-degrading enzymes and chemokine release. Following pro-inflammatory LPS activation, VLCFA levels increased also in healthy macrophages. With the onset of the resolution, VLCFAs were rapidly cleared in control macrophages by increased peroxisomal VLCFA degradation through liver-X-receptor mediated upregulation of ABCD1. ABCD1 deficiency impaired VLCFA homeostasis and prolonged pro-inflammatory gene expression upon LPS treatment. Our study uncovers a pivotal role for ABCD1, a protein linked to neuroinflammation, and associated peroxisomal VLCFA degradation in regulating macrophage plasticity.
Keywords: Extracellular matrix degradation; Immune response; Lipid metabolism; Neuroinflammation; X-linked adrenoleukodystrophy.
© 2022. The Author(s).
Conflict of interest statement
The authors have no competing interests.
Figures





References
-
- Turk BR, Theda C, Fatemi A, Moser AB. X-linked adrenoleukodystrophy: pathology, pathophysiology, diagnostic testing, newborn screening, and therapies. Int J Dev Neurosci. 2019;25:S0736-5748(19)30133-9. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
