

Endothelial cells regulate astrocyte to neural progenitor cell trans-differentiation in a mouse model of stroke
- PMID: 36535938
- PMCID: PMC9763251
- DOI: 10.1038/s41467-022-35498-6
Endothelial cells regulate astrocyte to neural progenitor cell trans-differentiation in a mouse model of stroke
Abstract
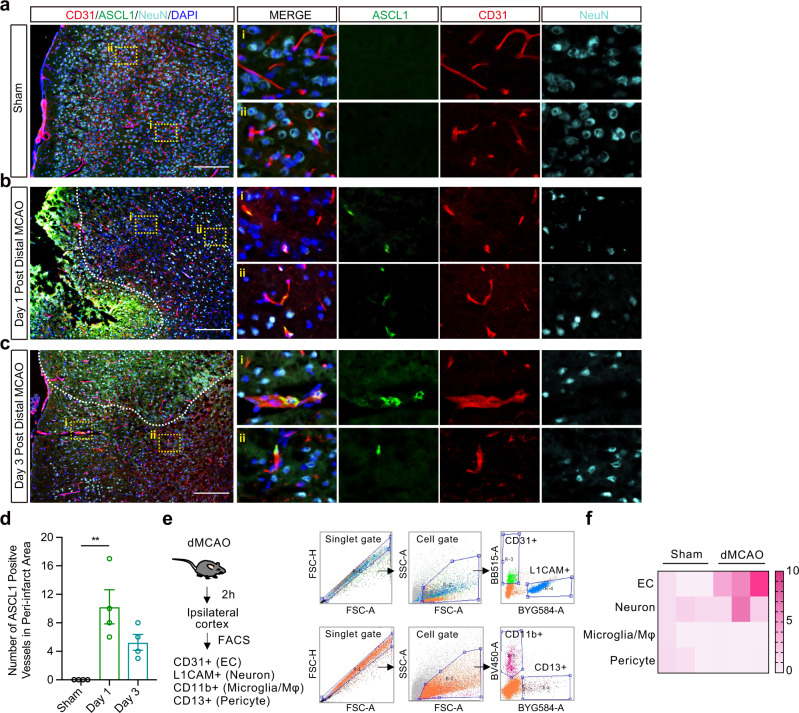
The concept of the neurovascular unit emphasizes the importance of cell-cell signaling between neural, glial, and vascular compartments. In neurogenesis, for example, brain endothelial cells play a key role by supplying trophic support to neural progenitors. Here, we describe a surprising phenomenon where brain endothelial cells may release trans-differentiation signals that convert astrocytes into neural progenitor cells in male mice after stroke. After oxygen-glucose deprivation, brain endothelial cells release microvesicles containing pro-neural factor Ascl1 that enter into astrocytes to induce their trans-differentiation into neural progenitors. In mouse models of focal cerebral ischemia, Ascl1 is upregulated in endothelium prior to astrocytic conversion into neural progenitor cells. Injecting brain endothelial-derived microvesicles amplifies the process of astrocyte trans-differentiation. Endothelial-specific overexpression of Ascl1 increases the local conversion of astrocytes into neural progenitors and improves behavioral recovery. Our findings describe an unexpected vascular-regulated mechanism of neuroplasticity that may open up therapeutic opportunities for improving outcomes after stroke.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
