

Plant microbiomes harbor potential to promote nutrient turnover in impoverished substrates of a Brazilian biodiversity hotspot
- PMID: 36536072
- PMCID: PMC9938248
- DOI: 10.1038/s41396-022-01345-1
Plant microbiomes harbor potential to promote nutrient turnover in impoverished substrates of a Brazilian biodiversity hotspot
Abstract
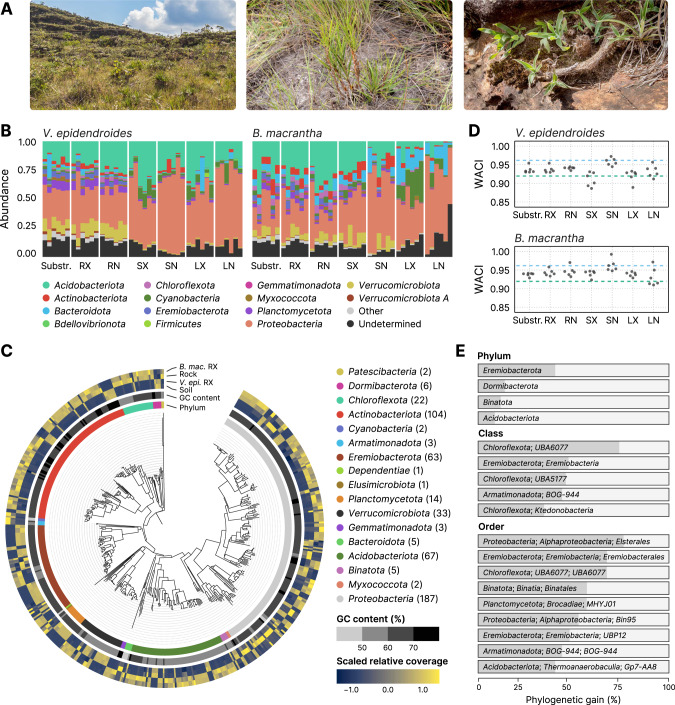
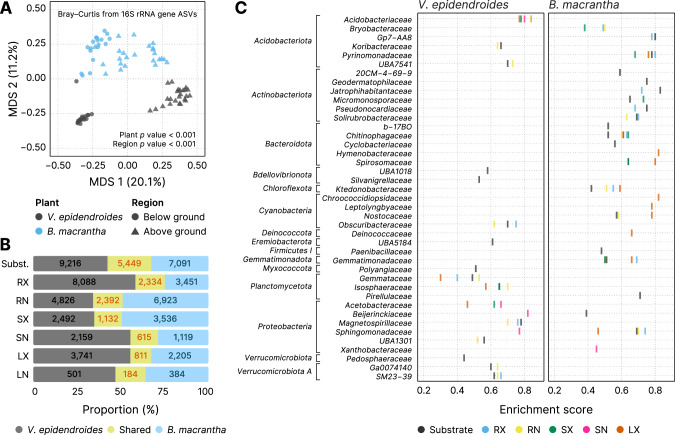
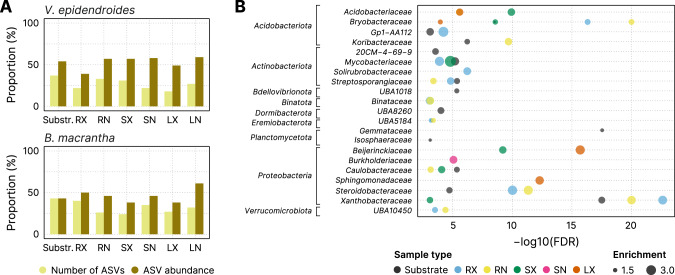
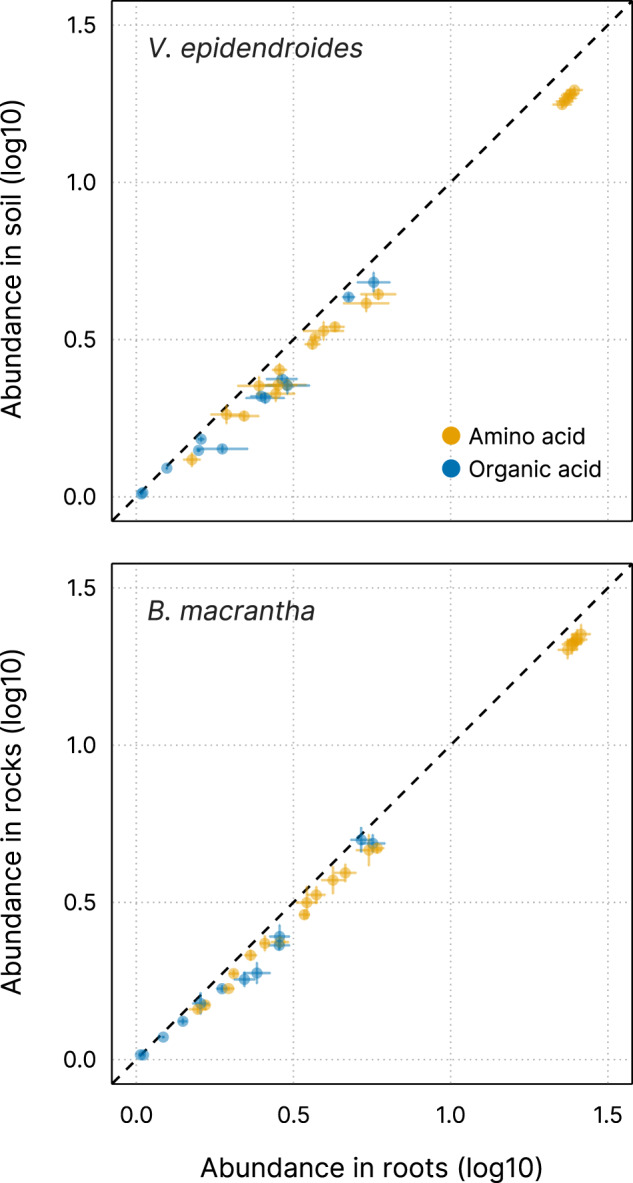
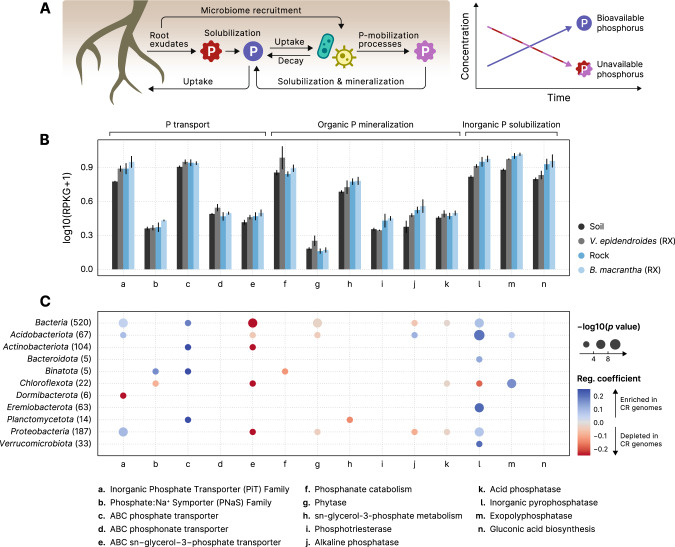
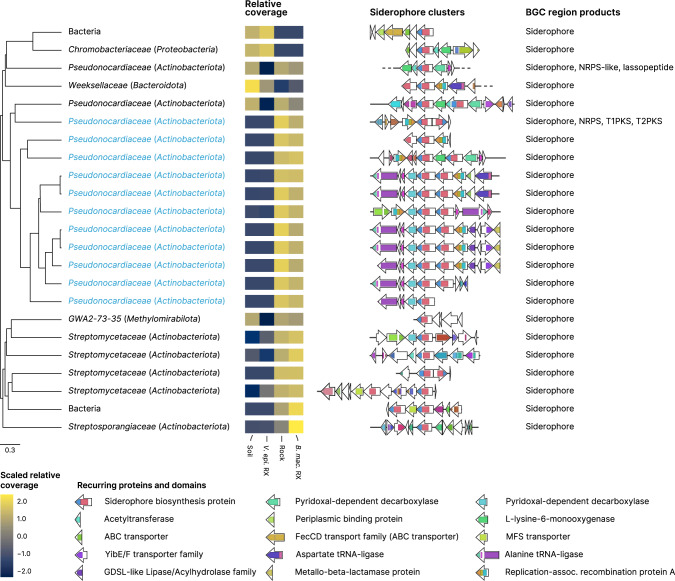
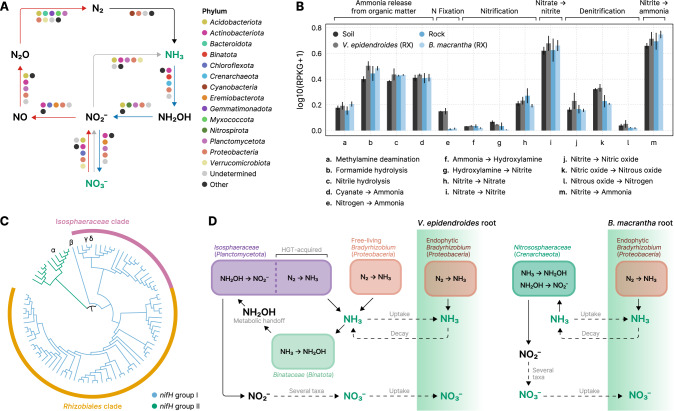
The substrates of the Brazilian campos rupestres, a grassland ecosystem, have extremely low concentrations of phosphorus and nitrogen, imposing restrictions to plant growth. Despite that, this ecosystem harbors almost 15% of the Brazilian plant diversity, raising the question of how plants acquire nutrients in such a harsh environment. Here, we set out to uncover the taxonomic profile, the compositional and functional differences and similarities, and the nutrient turnover potential of microbial communities associated with two plant species of the campos rupestres-dominant family Velloziaceae that grow over distinct substrates (soil and rock). Using amplicon sequencing data, we show that, despite the pronounced composition differentiation, the plant-associated soil and rock communities share a core of highly efficient colonizers that tend to be highly abundant and is enriched in 21 bacterial families. Functional investigation of metagenomes and 522 metagenome-assembled genomes revealed that the microorganisms found associated to plant roots are enriched in genes involved in organic compound intake, and phosphorus and nitrogen turnover. We show that potential for phosphorus transport, mineralization, and solubilization are mostly found within bacterial families of the shared microbiome, such as Xanthobacteraceae and Bryobacteraceae. We also detected the full repertoire of nitrogen cycle-related genes and discovered a lineage of Isosphaeraceae that acquired nitrogen-fixing potential via horizontal gene transfer and might be also involved in nitrification via a metabolic handoff association with Binataceae. We highlight that plant-associated microbial populations in the campos rupestres harbor a genetic repertoire with potential to increase nutrient availability and that the microbiomes of biodiversity hotspots can reveal novel mechanisms of nutrient turnover.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Oliveira RS, Abrahão A, Pereira C, Teodoro GS, Brum M, Alcantara S, et al. Ecophysiology of Campos Rupestres Plants. In: Fernandes GW, et al., editors. Ecology and Conservation of Mountaintop grasslands in Brazil. Cham: Springer International Publishing; 2016. pp. 227–72.
-
- Schaefer CEGR, Corrêa GR, Candido HG, Arruda DM, Nunes JA, Araujo RW, et al. The Physical Environment of Rupestrian Grasslands (Campos Rupestres) in Brazil: Geological, Geomorphological and Pedological Characteristics, and Interplays. In: Fernandes GW, et al., editors. Ecology and Conservation of Mountaintop grasslands in Brazil. Cham: Springer International Publishing; 2016. pp. 15–53.
-
- Silveira FAO, Negreiros D, Barbosa NPU, Buisson E, Carmo FF, Carstensen DW, et al. Ecology and evolution of plant diversity in the endangered campo rupestre: a neglected conservation priority. Plant Soil. 2016;403:129–52. doi: 10.1007/s11104-015-2637-8. - DOI
-
- Rapini A, Bitencourt C, Luebert F, Cardoso D. An escape-to-radiate model for explaining the high plant diversity and endemism in campos rupestres. Biol J Linn Soc. 2021;133:481–98. doi: 10.1093/biolinnean/blaa179. - DOI
-
- Negreiros D, Le Stradic S, Fernandes GW, Rennó HC. CSR analysis of plant functional types in highly diverse tropical grasslands of harsh environments. Plant Ecol. 2014;215:379–88. doi: 10.1007/s11258-014-0302-6. - DOI
