

A pioneer factor locally opens compacted chromatin to enable targeted ATP-dependent nucleosome remodeling
- PMID: 36536103
- PMCID: PMC10004348
- DOI: 10.1038/s41594-022-00886-5
A pioneer factor locally opens compacted chromatin to enable targeted ATP-dependent nucleosome remodeling
Abstract
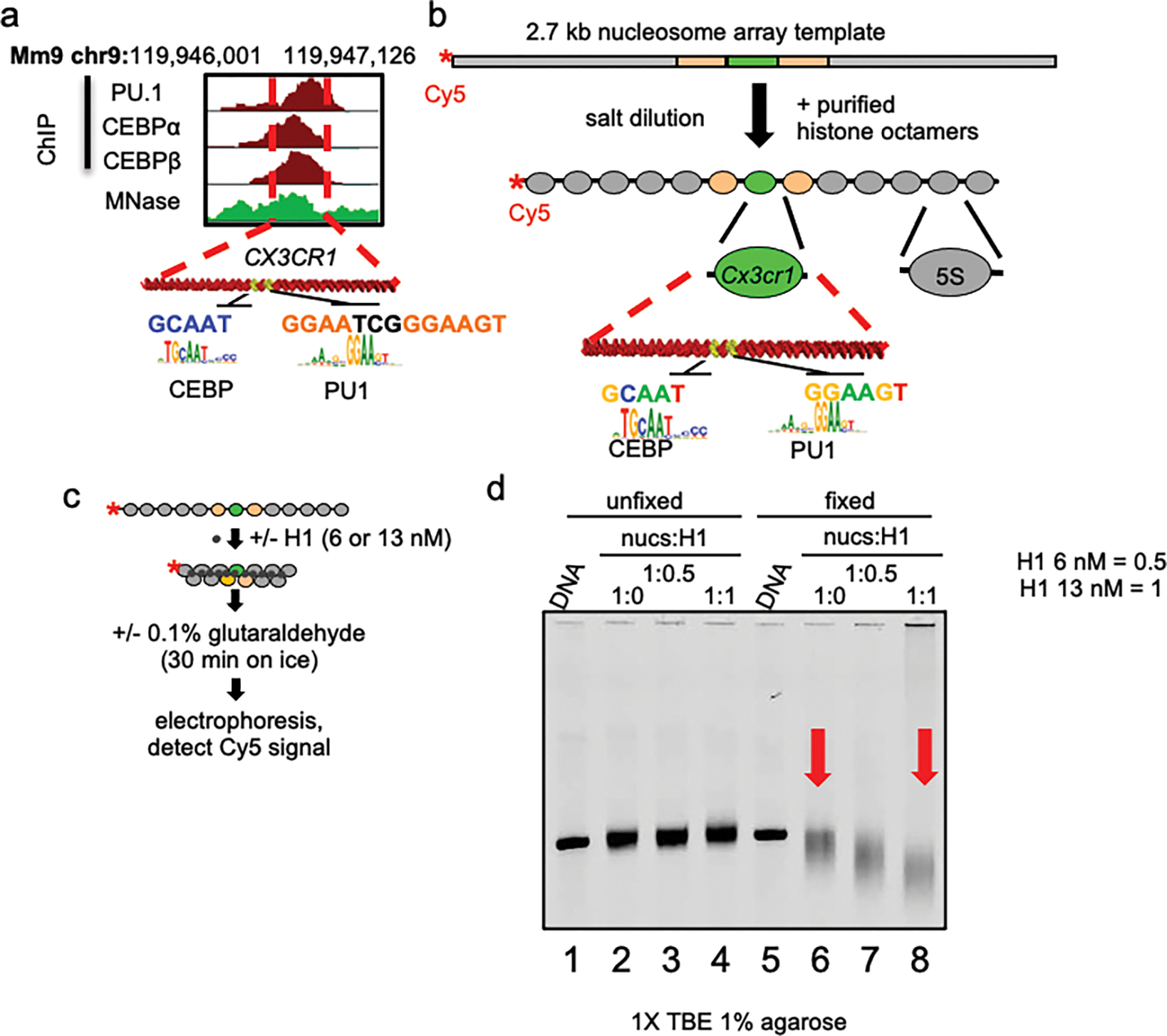
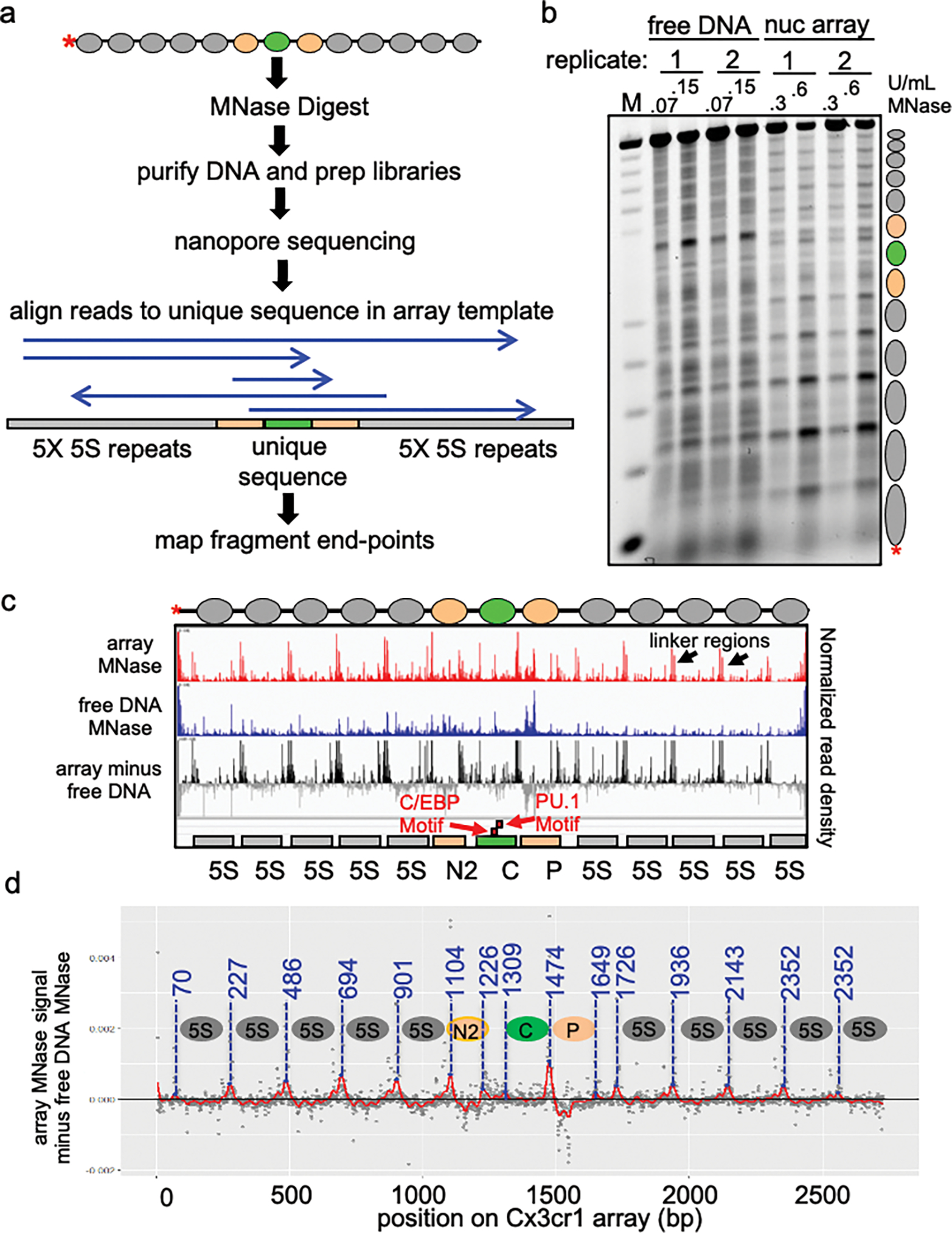
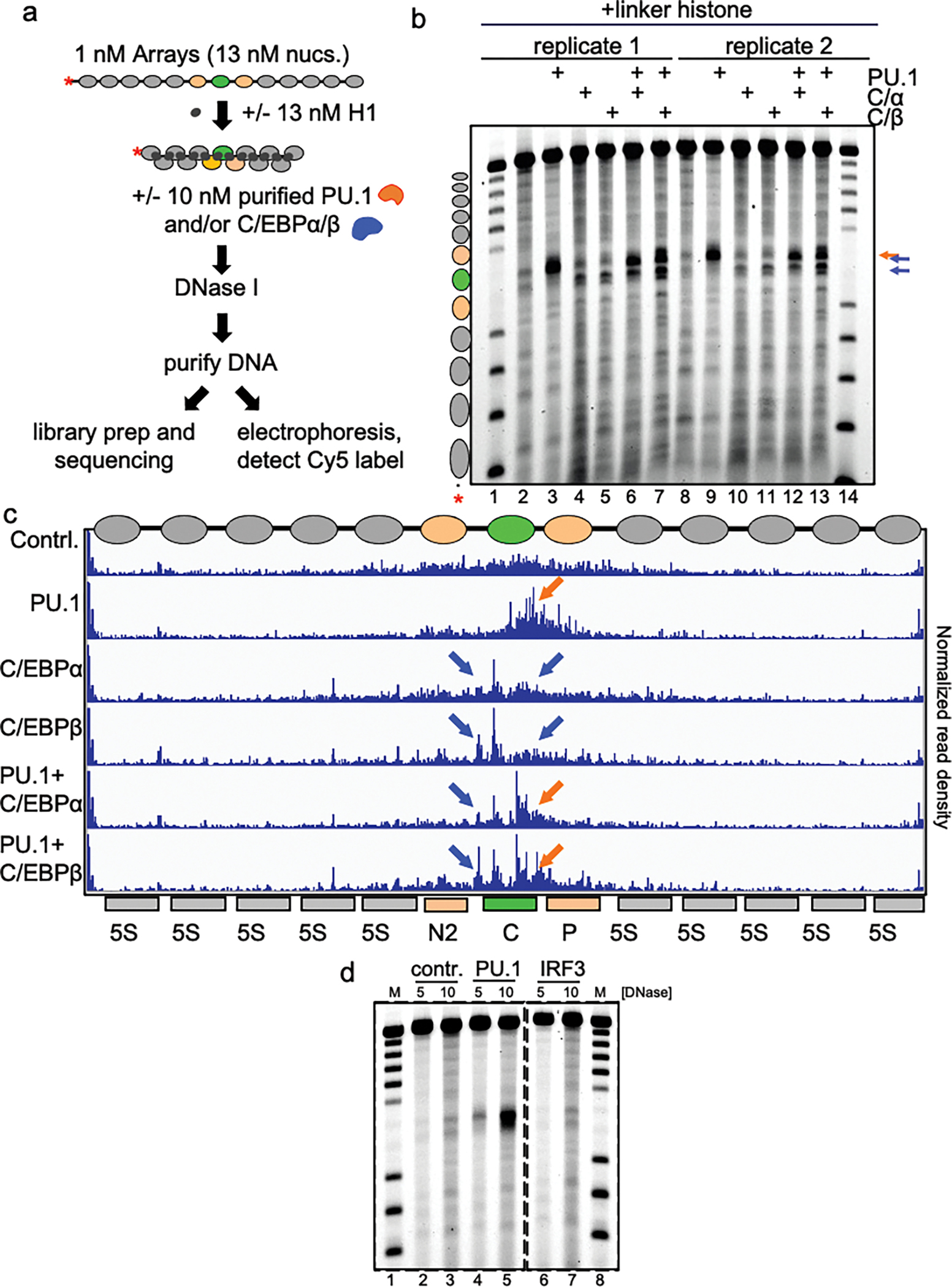
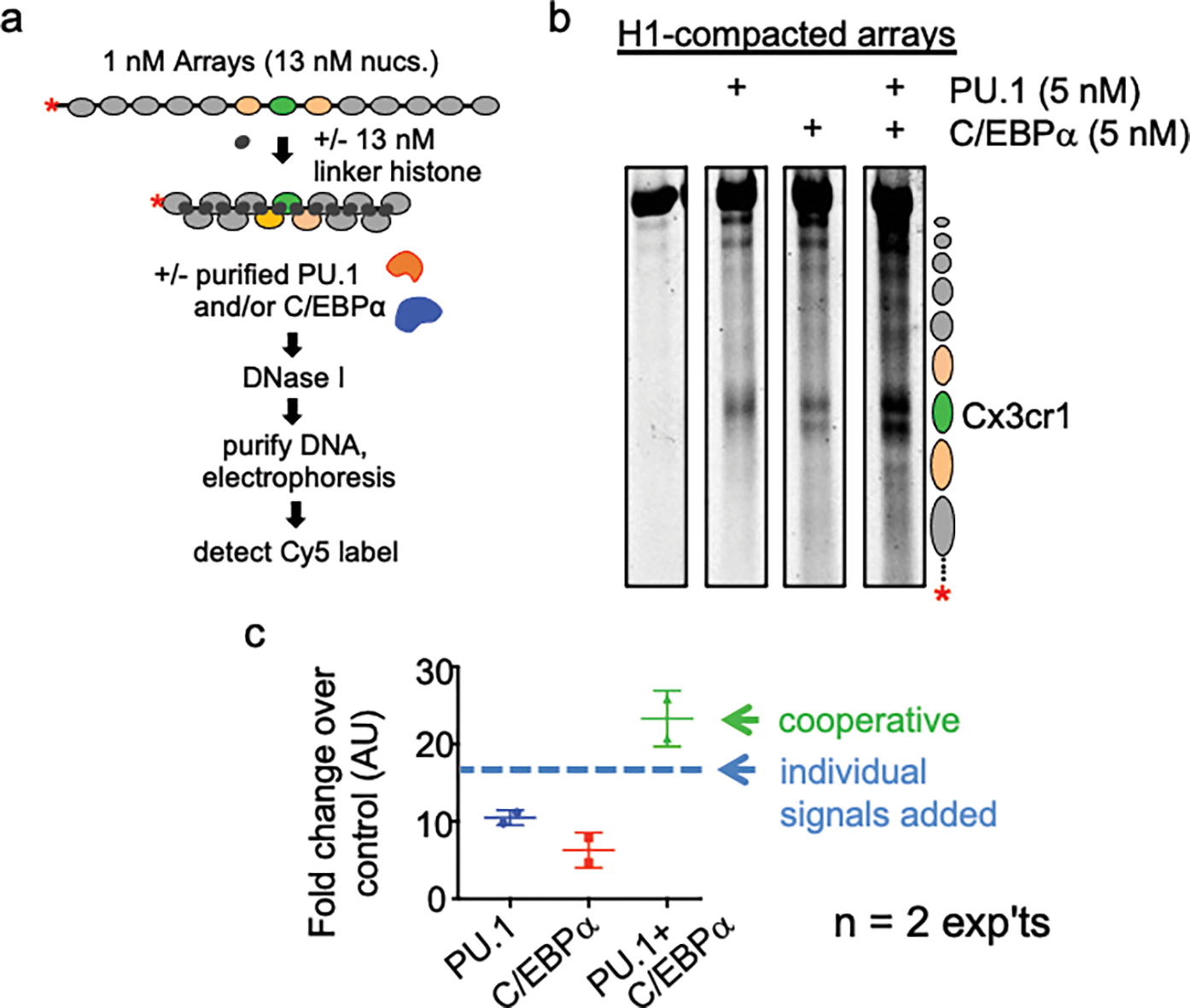
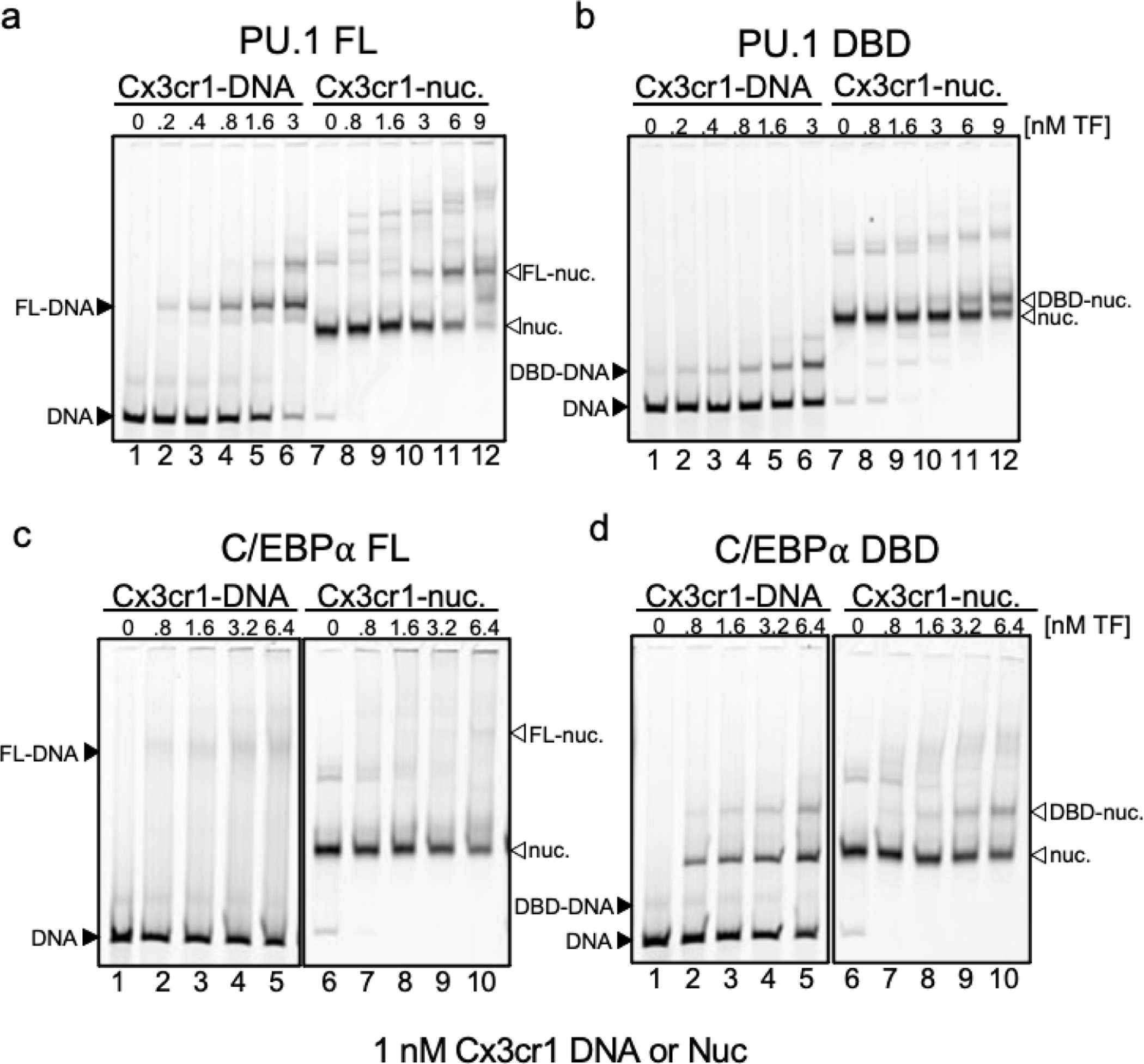
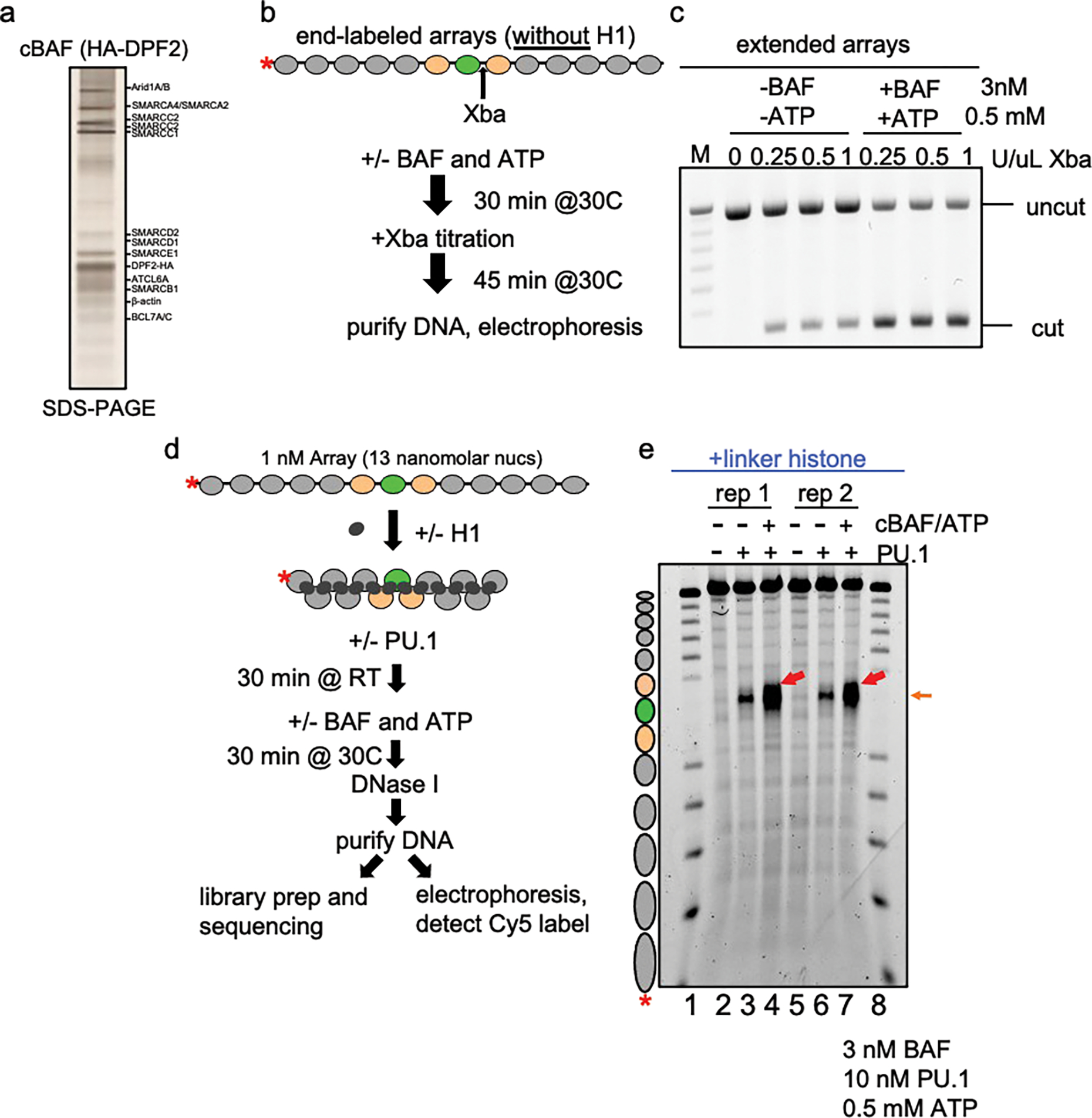
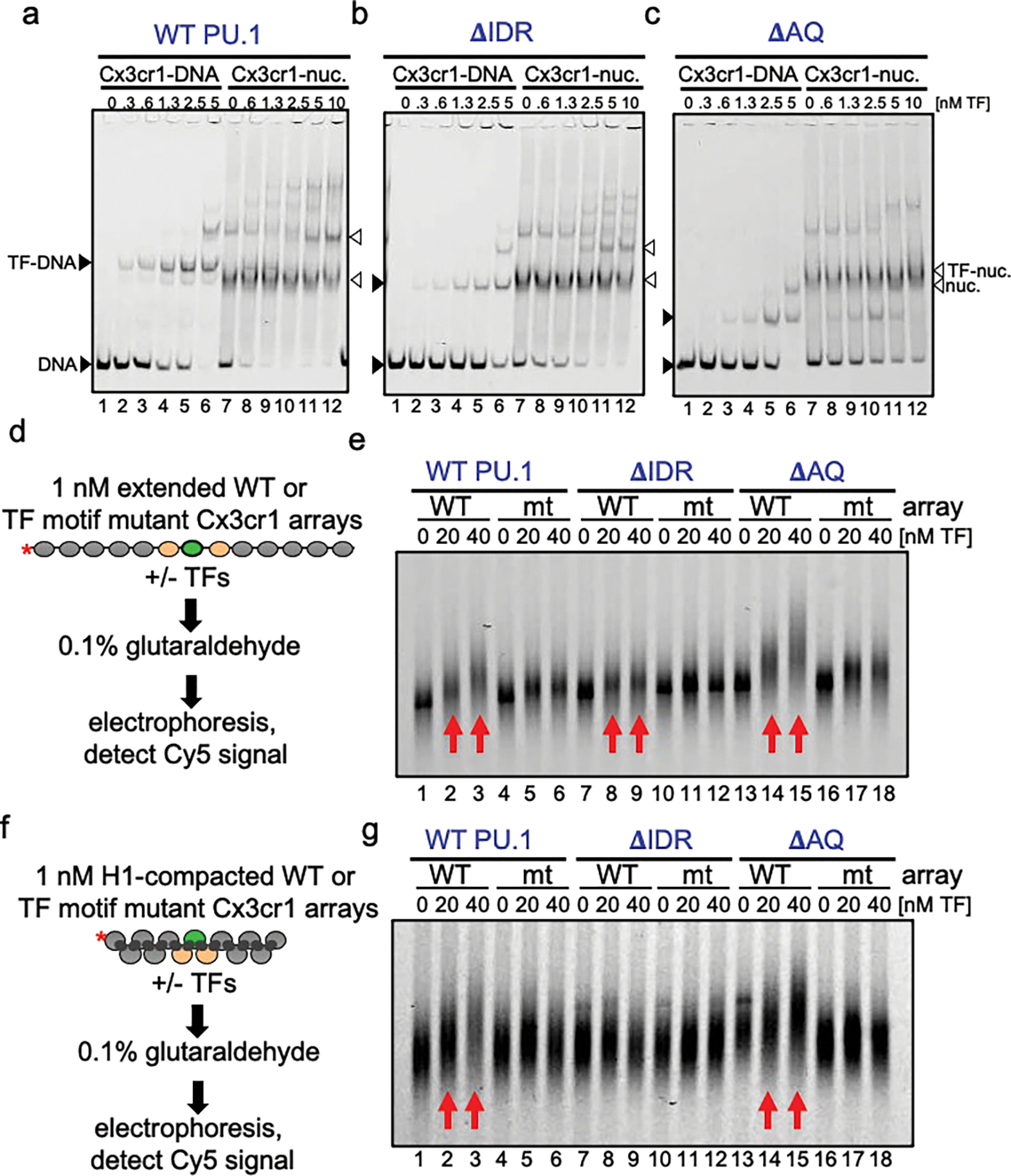
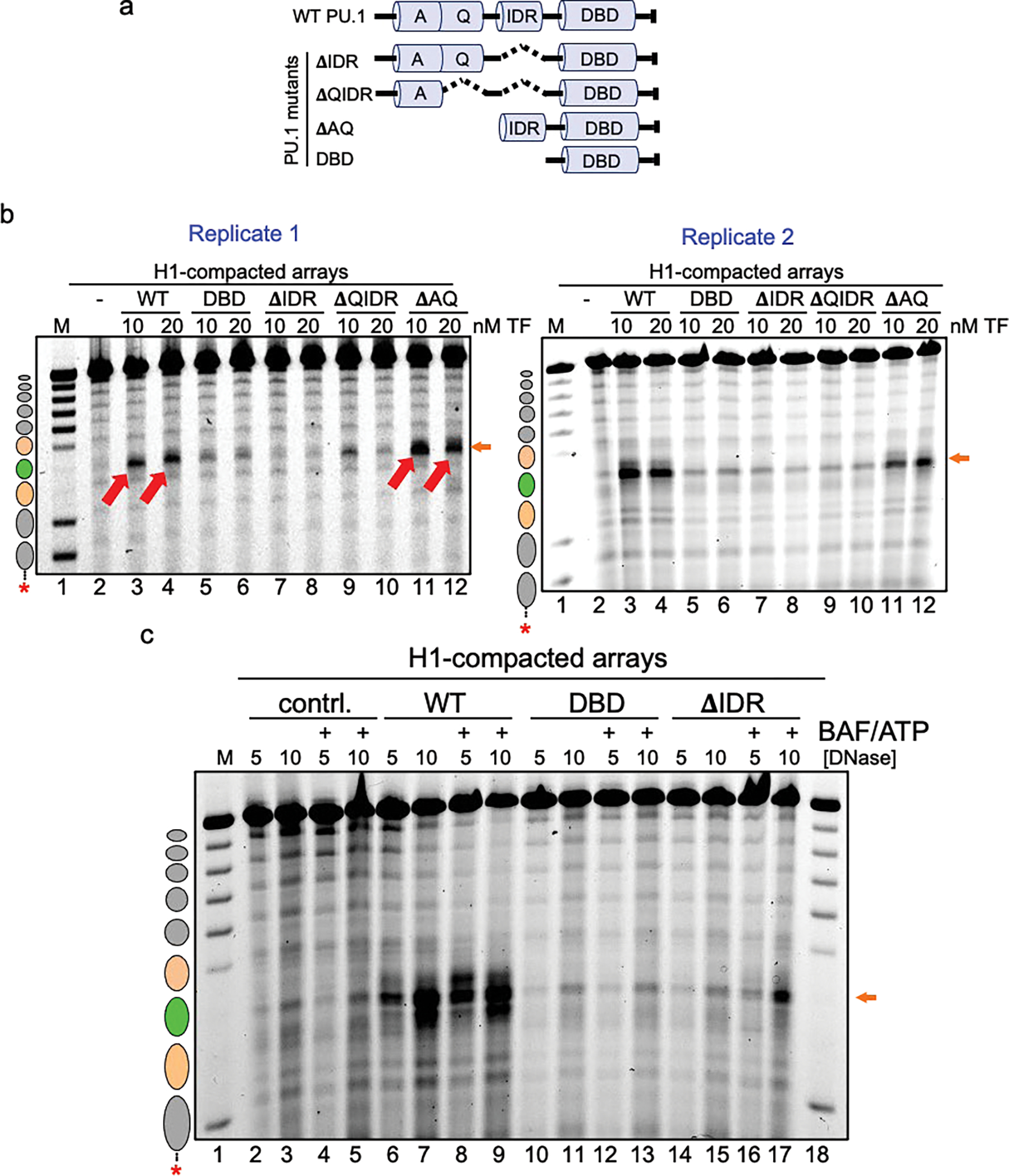
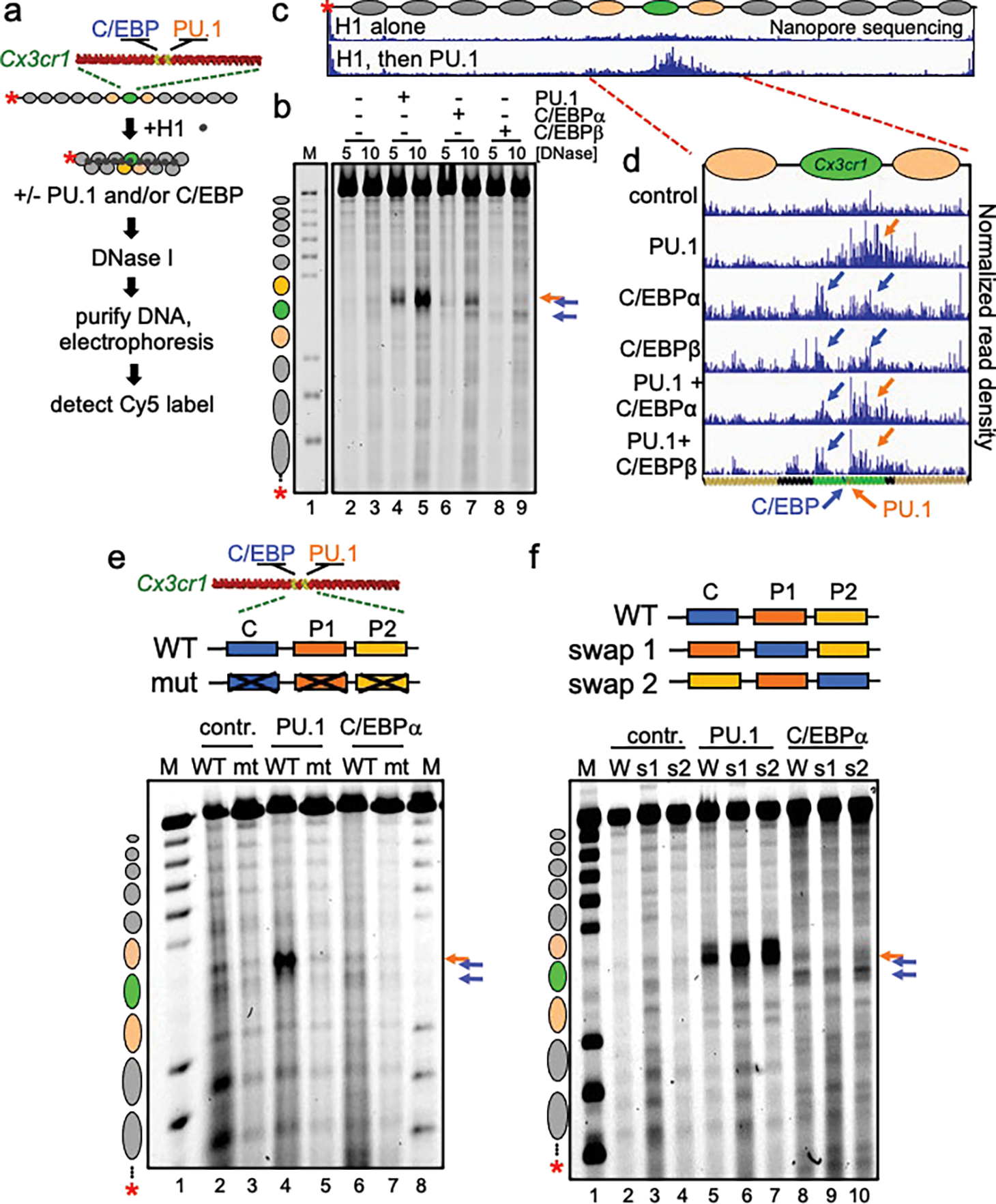
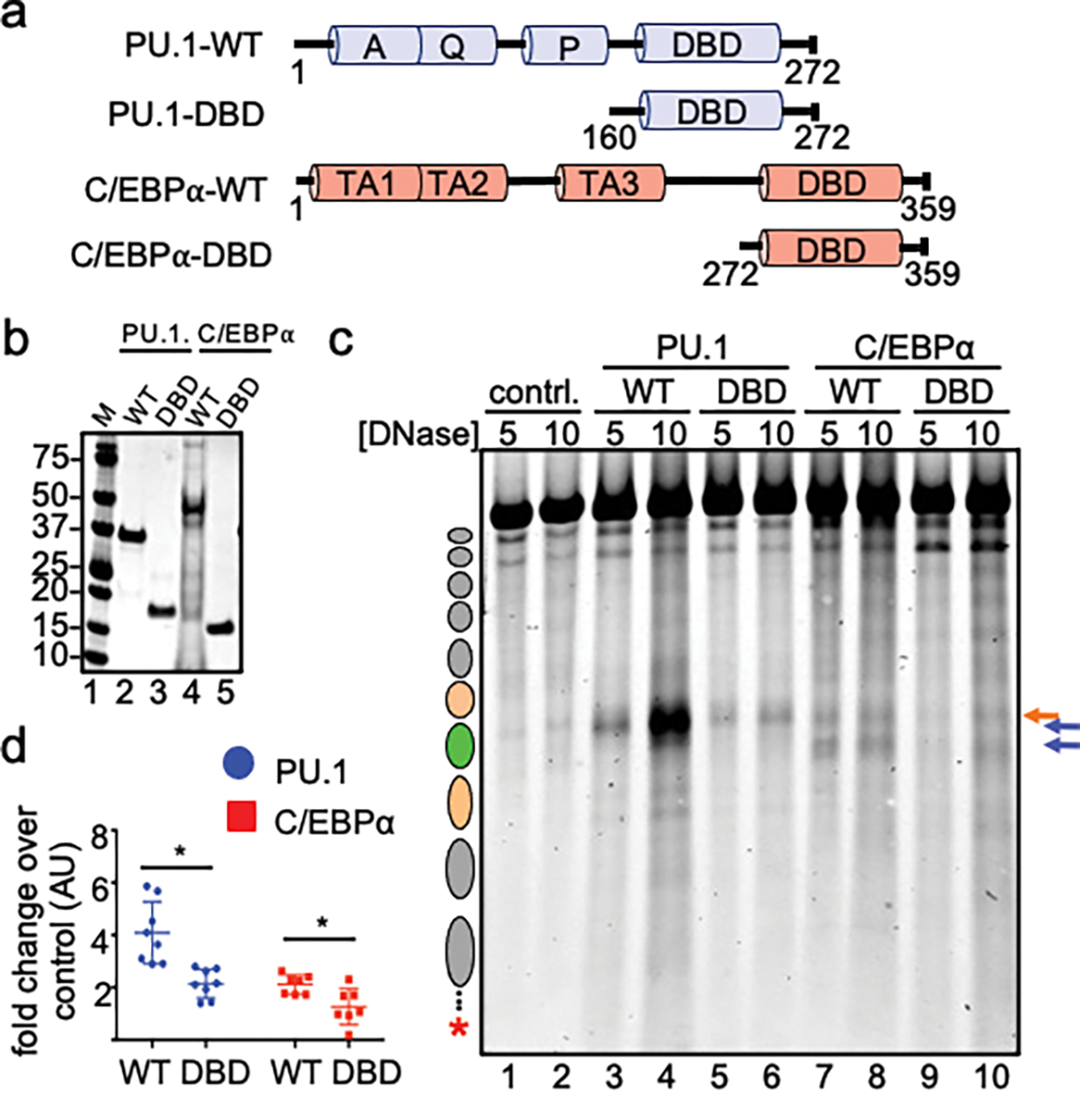
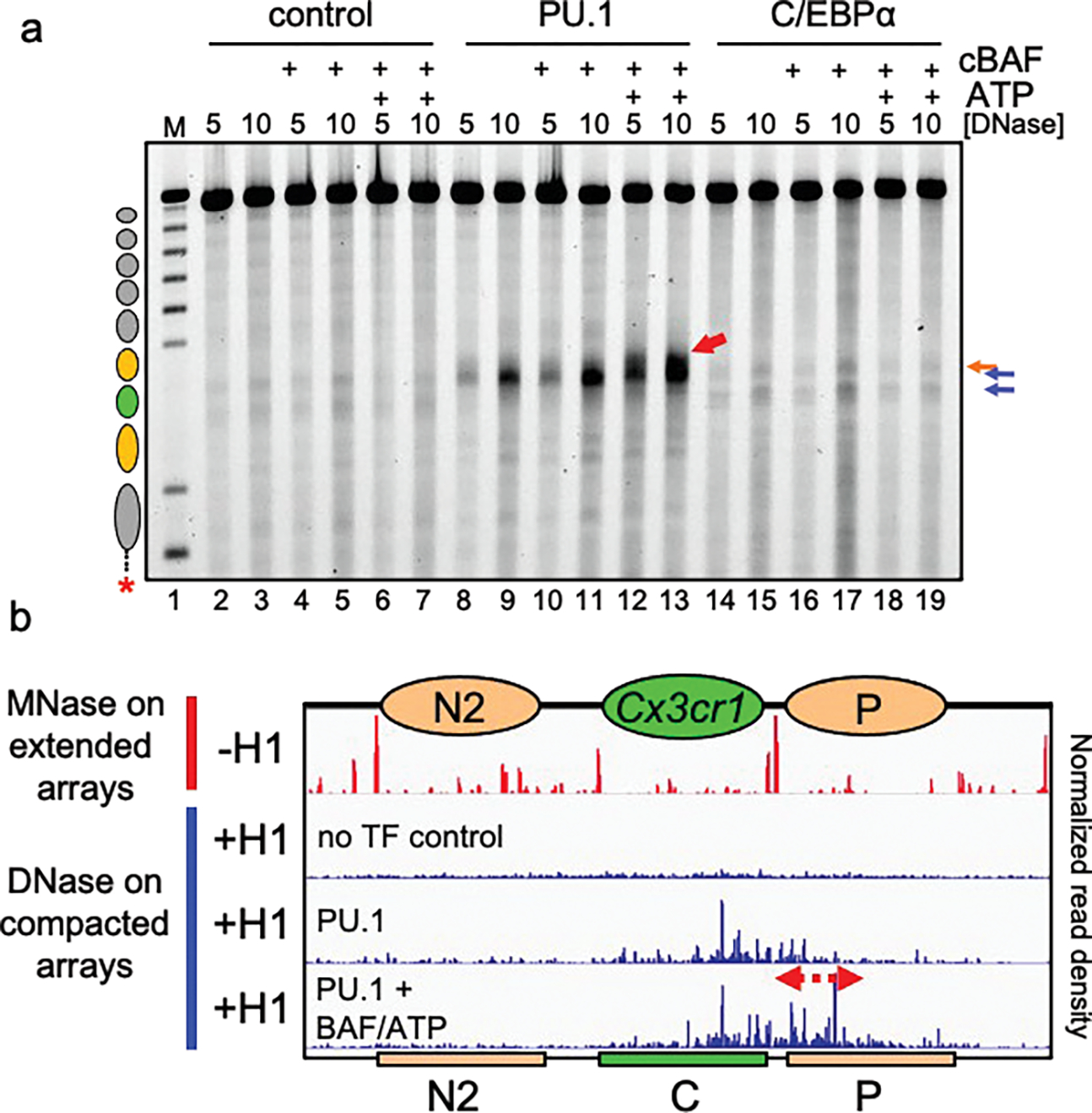
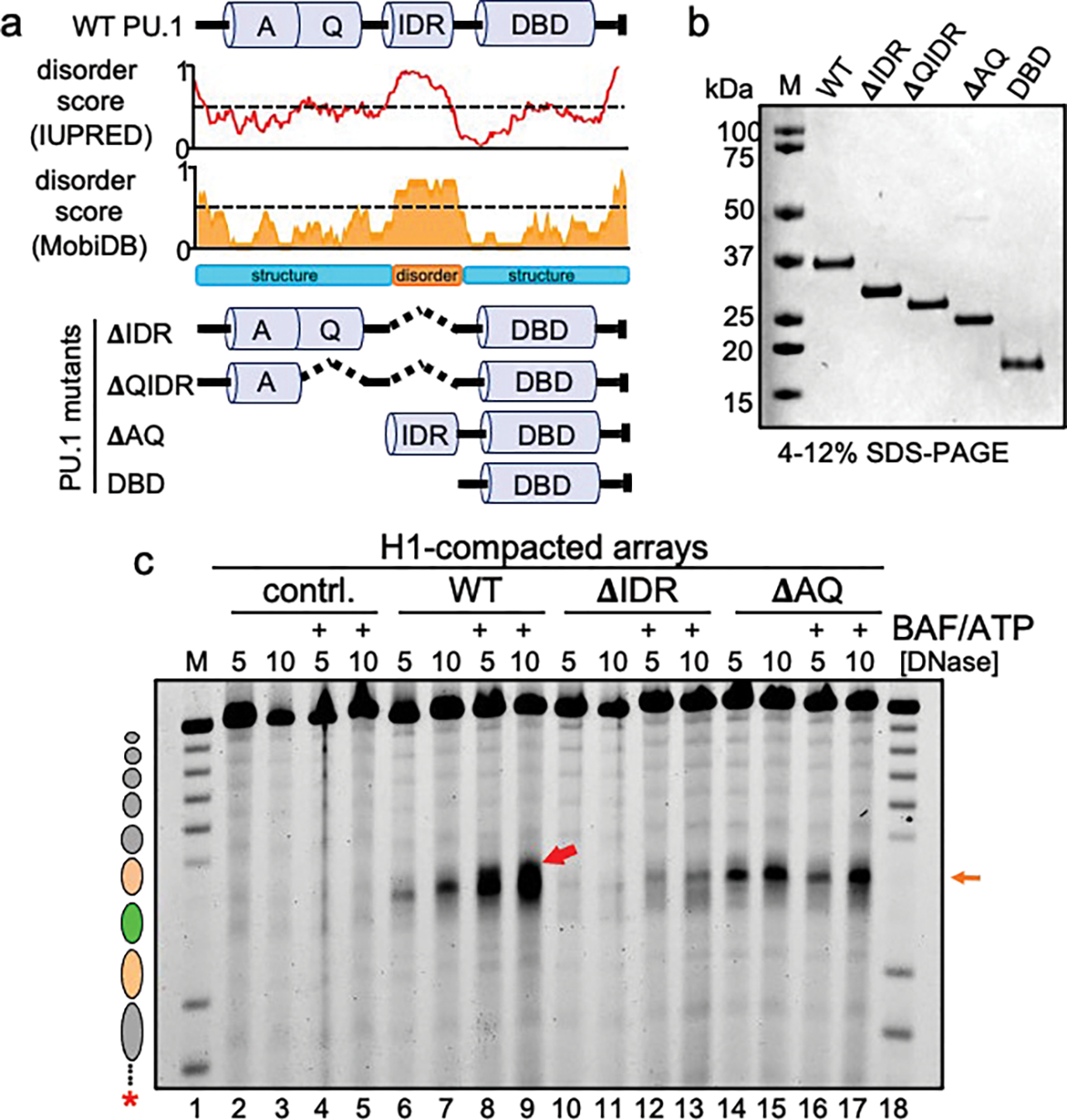
To determine how different pioneer transcription factors form a targeted, accessible nucleosome within compacted chromatin and collaborate with an ATP-dependent chromatin remodeler, we generated nucleosome arrays in vitro with a central nucleosome containing binding sites for the hematopoietic E-Twenty Six (ETS) factor PU.1 and Basic Leucine Zipper (bZIP) factors C/EBPα and C/EBPβ. Our long-read sequencing reveals that each factor can expose a targeted nucleosome on linker histone-compacted arrays, but with different nuclease sensitivity patterns. The DNA binding domain of PU.1 binds mononucleosomes, but requires an additional intrinsically disordered domain to bind and open compacted chromatin. The canonical mammalian SWI/SNF (cBAF) remodeler was unable to act upon two forms of locally open chromatin unless cBAF was enabled by a separate transactivation domain of PU.1. cBAF potentiates the PU.1 DNA binding domain to weakly open chromatin in the absence of the PU.1 disordered domain. Our findings reveal a hierarchy by which chromatin is opened and show that pioneer factors can provide specificity for action by nucleosome remodelers.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Figures
References
-
- Schwarz PM & Hansen JC Formation and stability of higher order chromatin structures Contributions of the histone octamer. Journal of Biological Chemistry (1994). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
