

Polλ promotes microhomology-mediated end-joining
- PMID: 36536104
- PMCID: PMC10197178
- DOI: 10.1038/s41594-022-00895-4
Polλ promotes microhomology-mediated end-joining
Erratum in
-
Author Correction: Polλ promotes microhomology-mediated end-joining.Nat Struct Mol Biol. 2023 Jul;30(7):1048. doi: 10.1038/s41594-023-00993-x. Nat Struct Mol Biol. 2023. PMID: 37059901 No abstract available.
Abstract
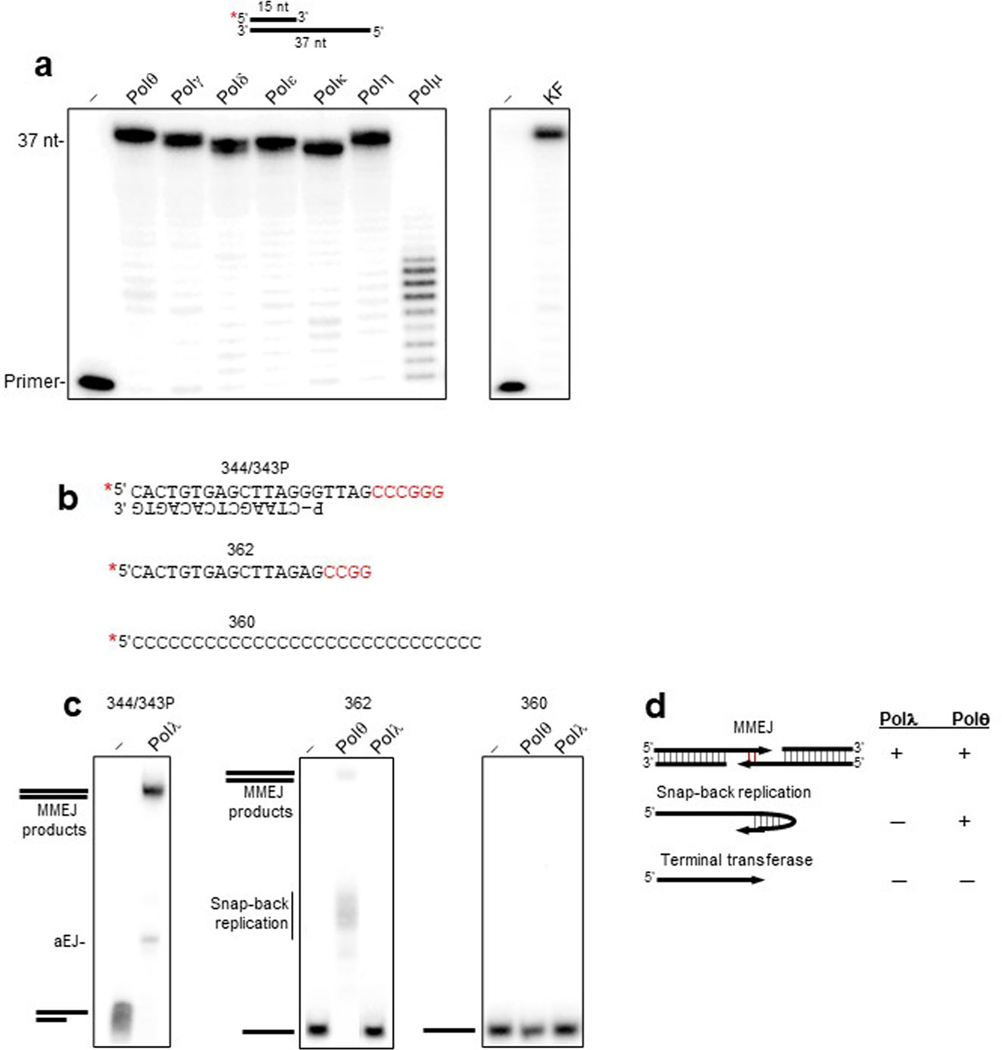
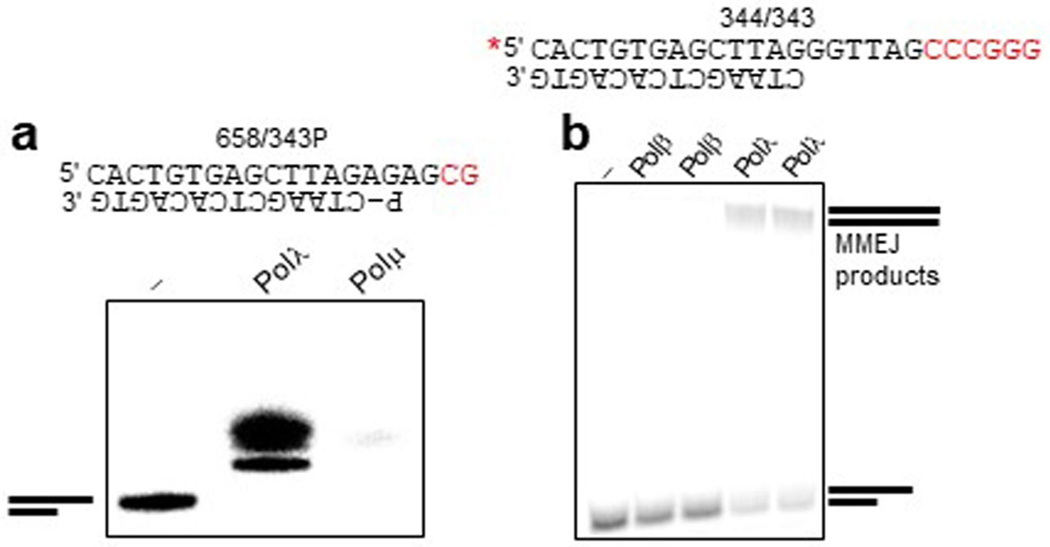
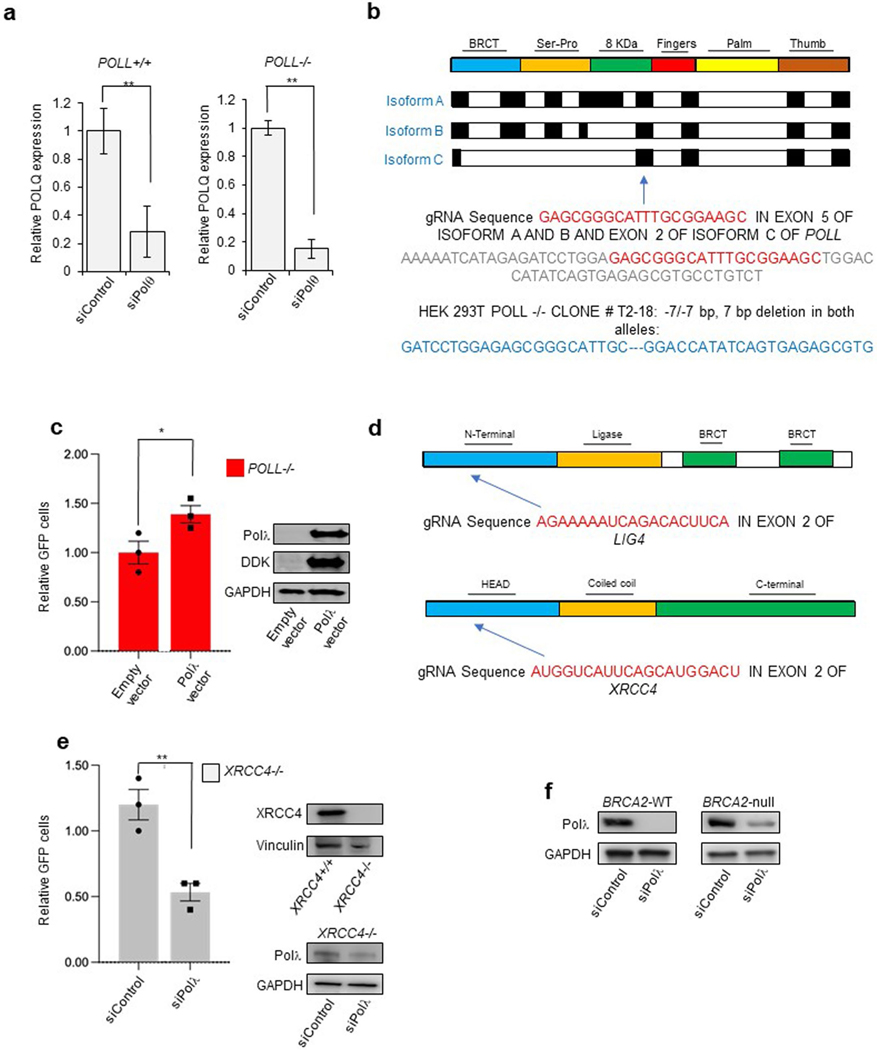
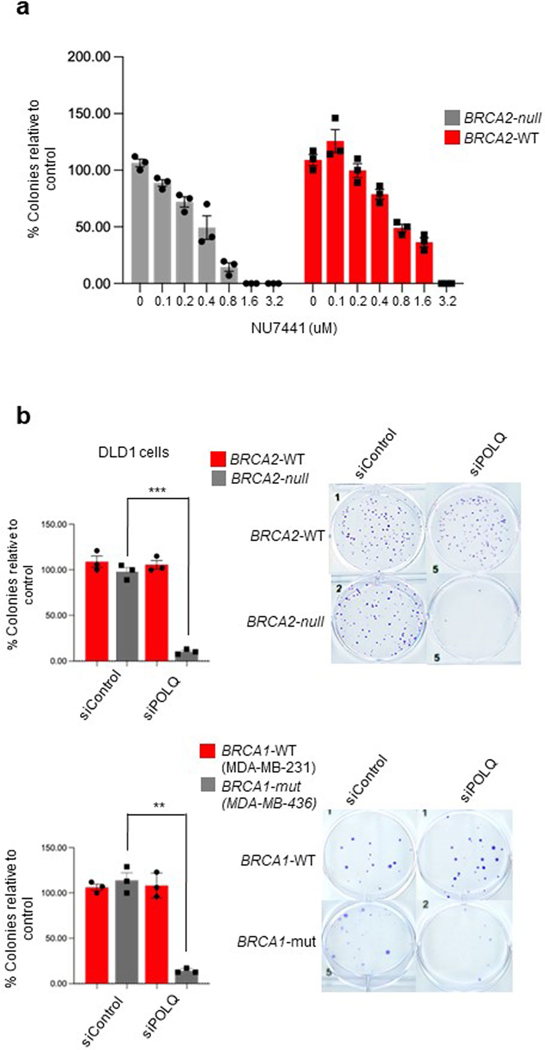
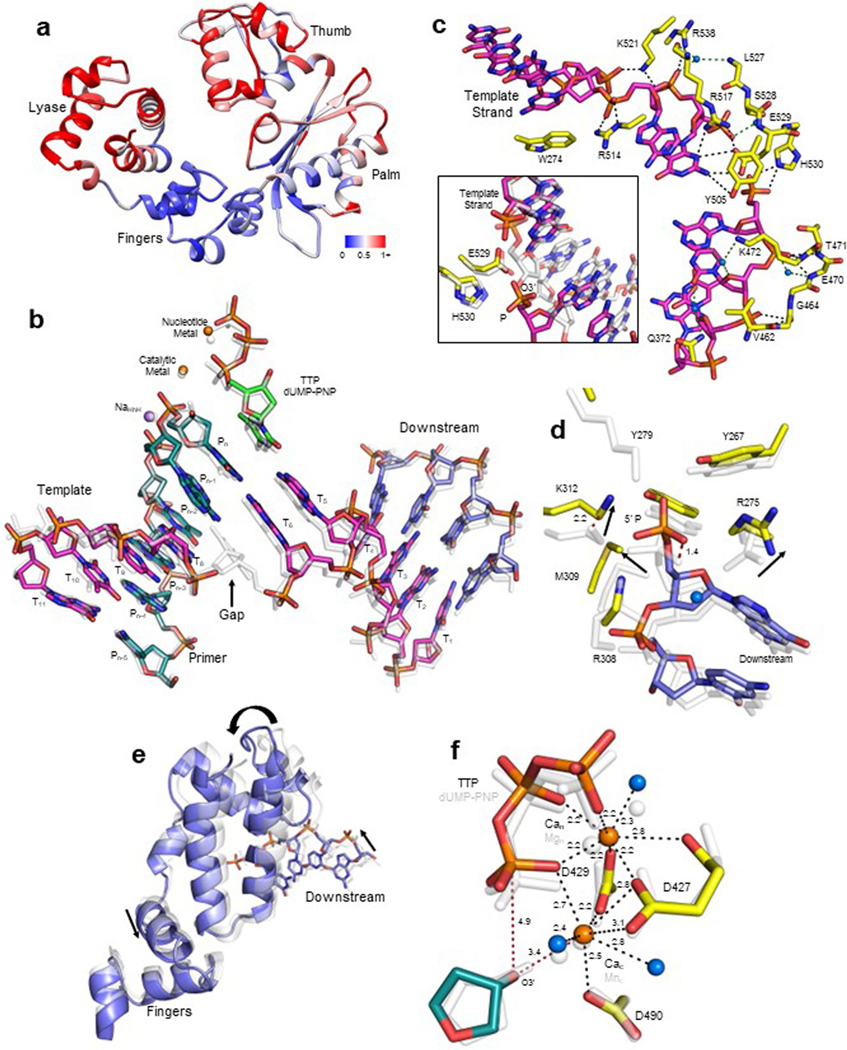
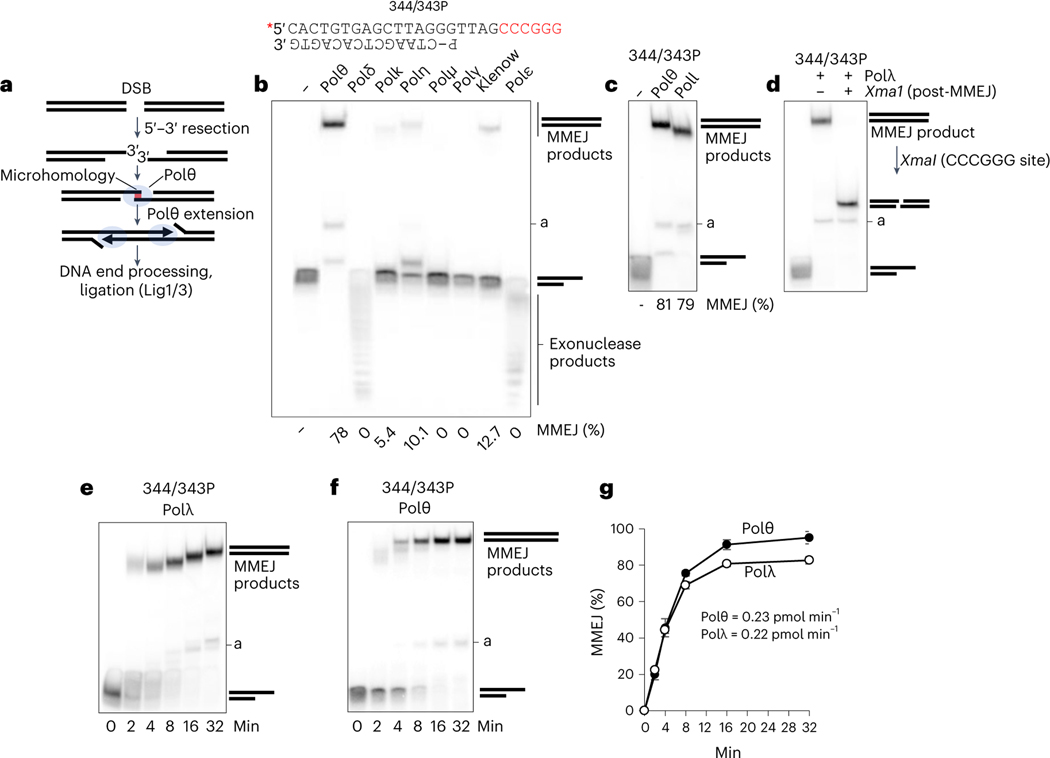
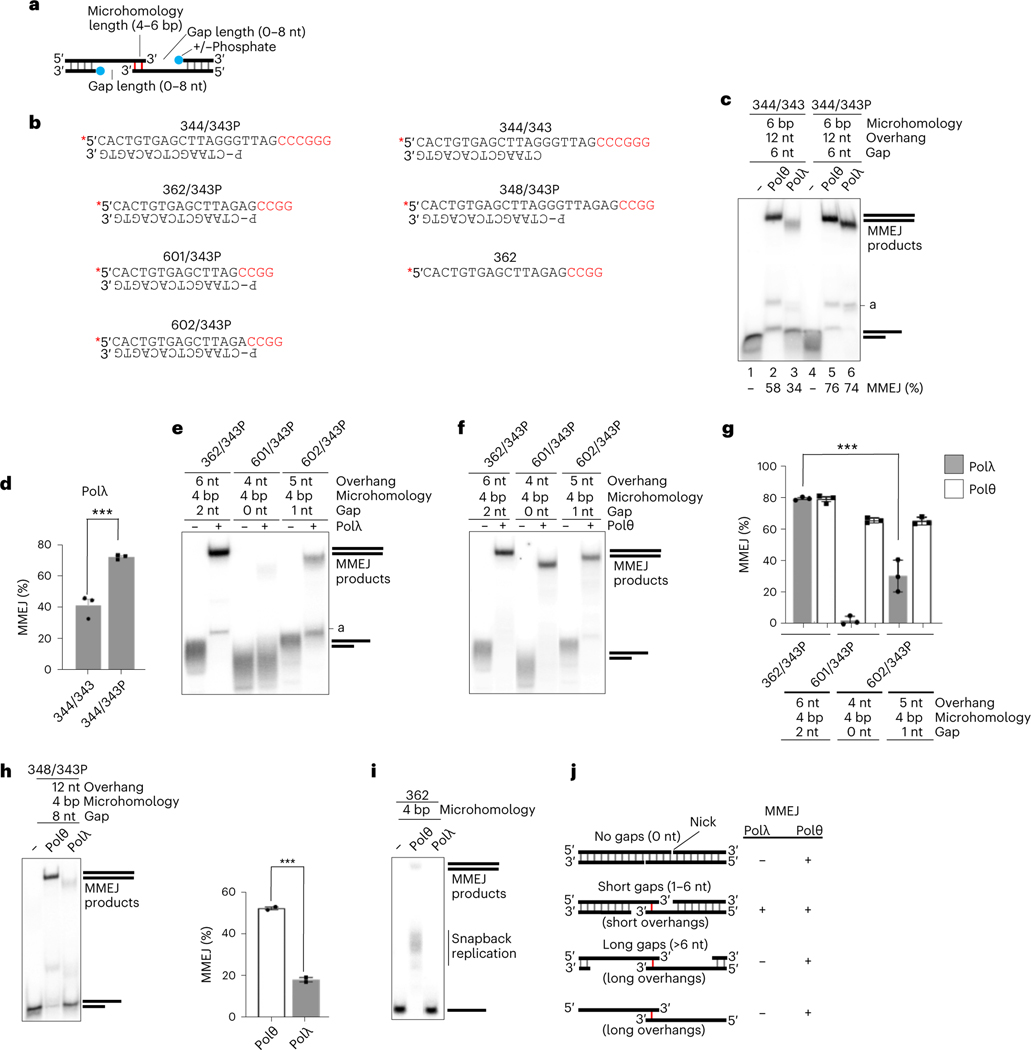
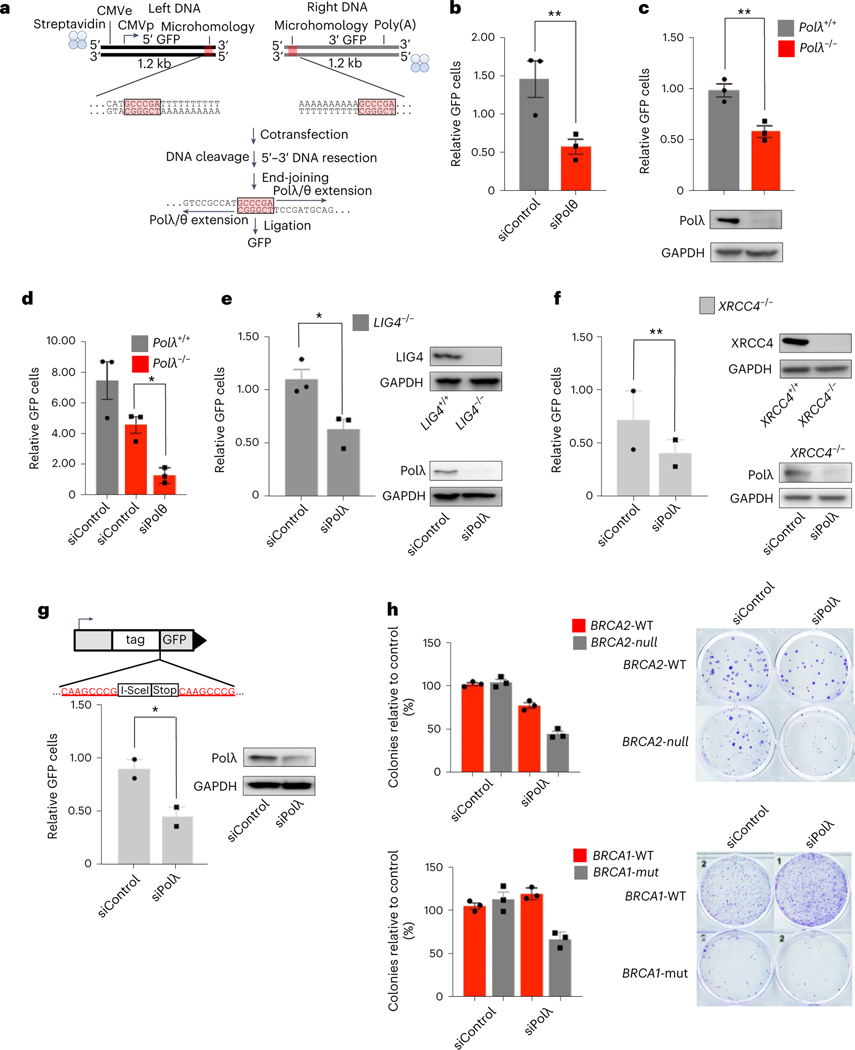
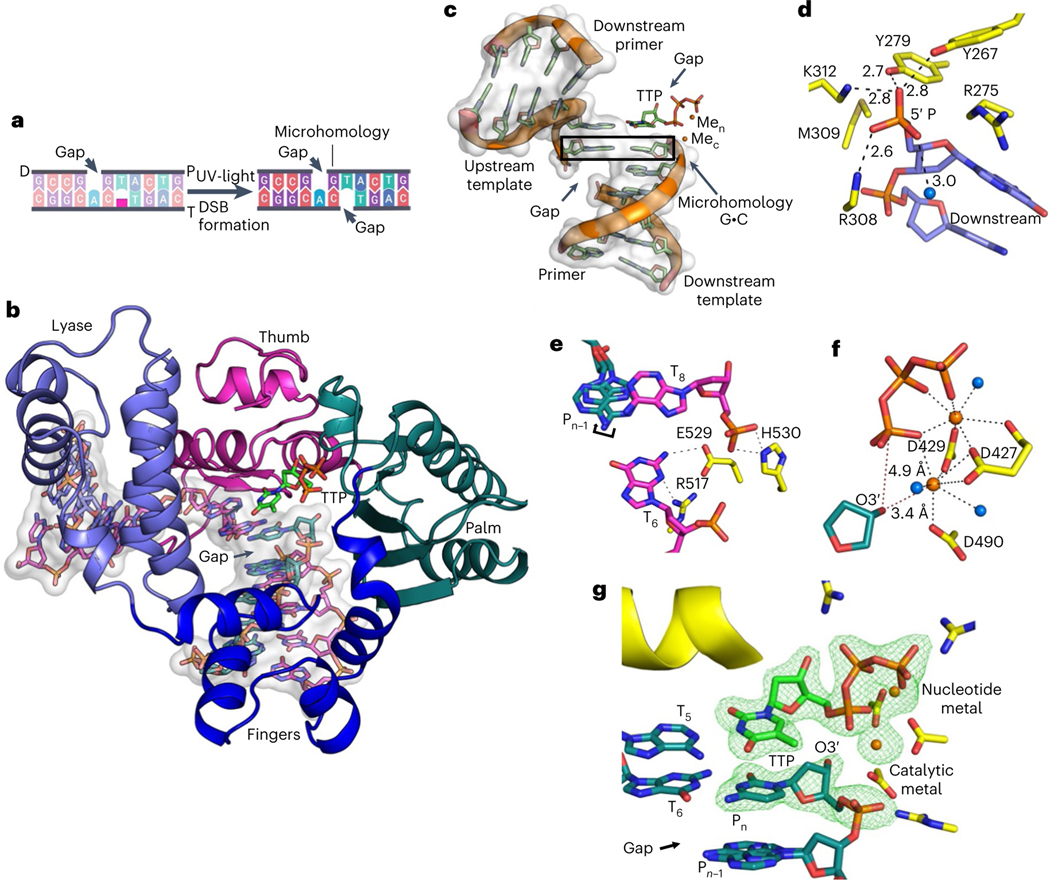
The double-strand break (DSB) repair pathway called microhomology-mediated end-joining (MMEJ) is thought to be dependent on DNA polymerase theta (Polθ) and occur independently of nonhomologous end-joining (NHEJ) factors. An unresolved question is whether MMEJ is facilitated by a single Polθ-mediated end-joining pathway or consists of additional undiscovered pathways. We find that human X-family Polλ, which functions in NHEJ, additionally exhibits robust MMEJ activity like Polθ. Polλ promotes MMEJ in mammalian cells independently of essential NHEJ factors LIG4/XRCC4 and Polθ, which reveals a distinct Polλ-dependent MMEJ mechanism. X-ray crystallography employing in situ photo-induced DSB formation captured Polλ in the act of stabilizing a microhomology-mediated DNA synapse with incoming nucleotide at 2.0 Å resolution and reveals how Polλ performs replication across a DNA synapse joined by minimal base-pairing. Last, we find that Polλ is semisynthetic lethal with BRCA1 and BRCA2. Together, these studies indicate Polλ MMEJ as a distinct DSB repair mechanism.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Figures
References
-
- Yan CT et al. IgH class switching and translocations use a robust non-classical end-joining pathway. Nature 449, 478–482 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
