

TGFβ+ small extracellular vesicles from head and neck squamous cell carcinoma cells reprogram macrophages towards a pro-angiogenic phenotype
- PMID: 36537293
- PMCID: PMC9764108
- DOI: 10.1002/jev2.12294
TGFβ+ small extracellular vesicles from head and neck squamous cell carcinoma cells reprogram macrophages towards a pro-angiogenic phenotype
Abstract
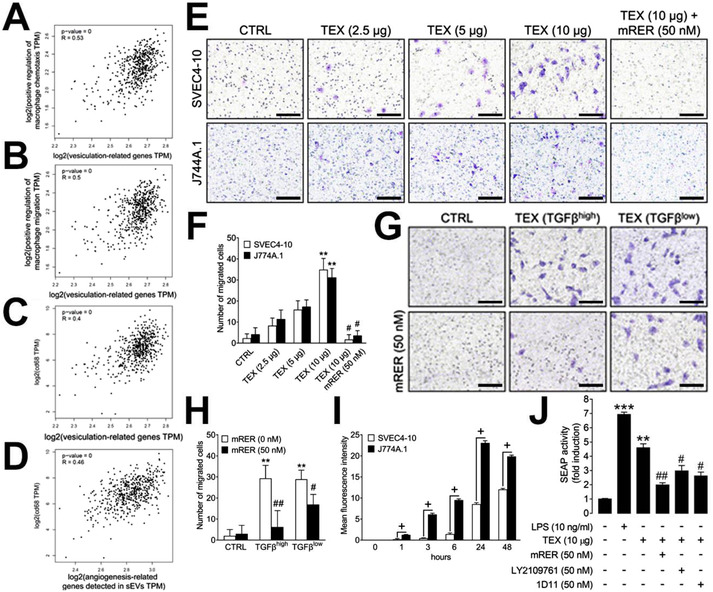
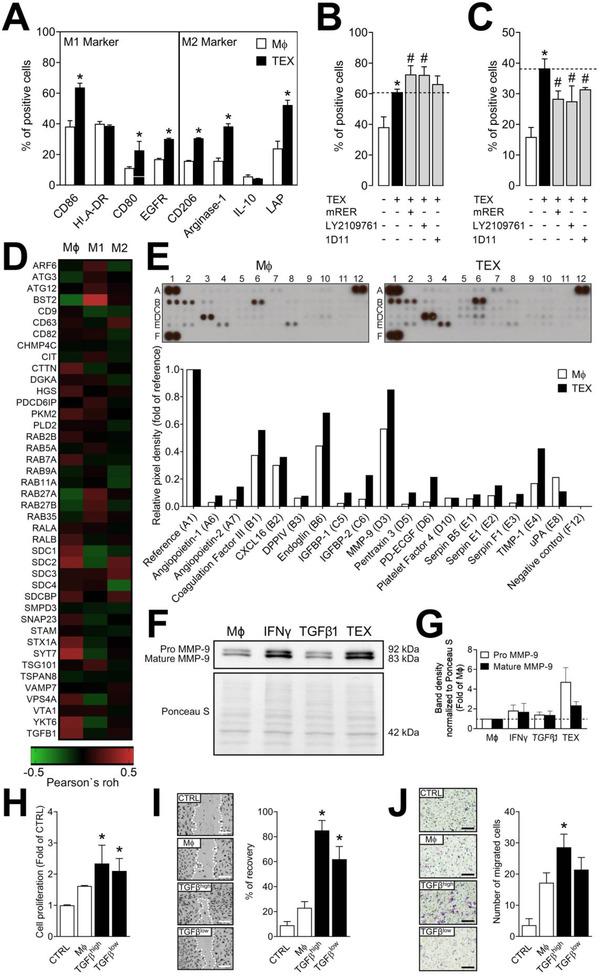
Transforming growth factor β (TGFβ) is a major component of tumor-derived small extracellular vesicles (TEX) in cancer patients. Mechanisms utilized by TGFβ+ TEX to promote tumor growth and pro-tumor activities in the tumor microenvironment (TME) are largely unknown. TEX produced by head and neck squamous cell carcinoma (HNSCC) cell lines carried TGFβ and angiogenesis-promoting proteins. TGFβ+ TEX stimulated macrophage chemotaxis without a notable M1/M2 phenotype shift and reprogrammed primary human macrophages to a pro-angiogenic phenotype characterized by the upregulation of pro-angiogenic factors and functions. In a murine basement membrane extract plug model, TGFβ+ TEX promoted macrophage infiltration and vascularization (p < 0.001), which was blocked by using the TGFβ ligand trap mRER (p < 0.001). TGFβ+ TEX injected into mice undergoing the 4-nitroquinoline-1-oxide (4-NQO)-driven oral carcinogenesis promoted tumor angiogenesis (p < 0.05), infiltration of M2-like macrophages in the TME (p < 0.05) and ultimately tumor progression (p < 0.05). Inhibition of TGFβ signaling in TEX with mRER ameliorated these pro-tumor activities. Silencing of TGFβ emerges as a critical step in suppressing pro-angiogenic functions of TEX in HNSCC.
Keywords: TGFβ; angiogenesis; exosomes; head and neck squamous cell carcinoma; macrophages; small extracellular vesicles.
© 2022 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles.
Conflict of interest statement
Dr. Andrew Hinck is the Co‐Inventor of RER, which is covered by U.S. patent 9,611,306, and holds royalty rights for the clinical deployment of RER, which is currently being pursued.
Figures




References
-
- Abnaof, K. , Mallela, N. , Walenda, G. , Meurer, S. K. , Seré, K. , Lin, Q. , Smeets, B. , Hoffmann, K. , Wagner, W. , Zenke, M. , Weiskirchen, R. , & Fröhlich, H. (2014). TGF‐β stimulation in human and murine cells reveals commonly affected biological processes and pathways at transcription level. BMC Systems Biology, 8, 1–14. - PMC - PubMed
-
- Bhatta, M. , Shenoy, G. N. , Loyall, J. L. , Gray, B. D. , Bapardekar, M. , Conway, A. , Minderman, H. , Kelleher Jr, R. J. , Carreno, B. M. , Linette, G. , Shultz, L. D. , Odunsi, K. , Balu‐Iyer, S. V. , Pak, K. Y. , & Bankert, R. B. (2021). Novel phosphatidylserine‐binding molecule enhances antitumor T‐cell responses by targeting immunosuppressive exosomes in human tumor microenvironments. Journal for ImmunoTherapy of Cancer, 9, 1–13. - PMC - PubMed
-
- Carmeliet, P. (2003). Angiogenesis in health and disease. Nature Medicine, 9, 653–660. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
