

Microbiota-mediated colonization resistance: mechanisms and regulation
- PMID: 36539611
- PMCID: PMC10249723
- DOI: 10.1038/s41579-022-00833-7
Microbiota-mediated colonization resistance: mechanisms and regulation
Abstract
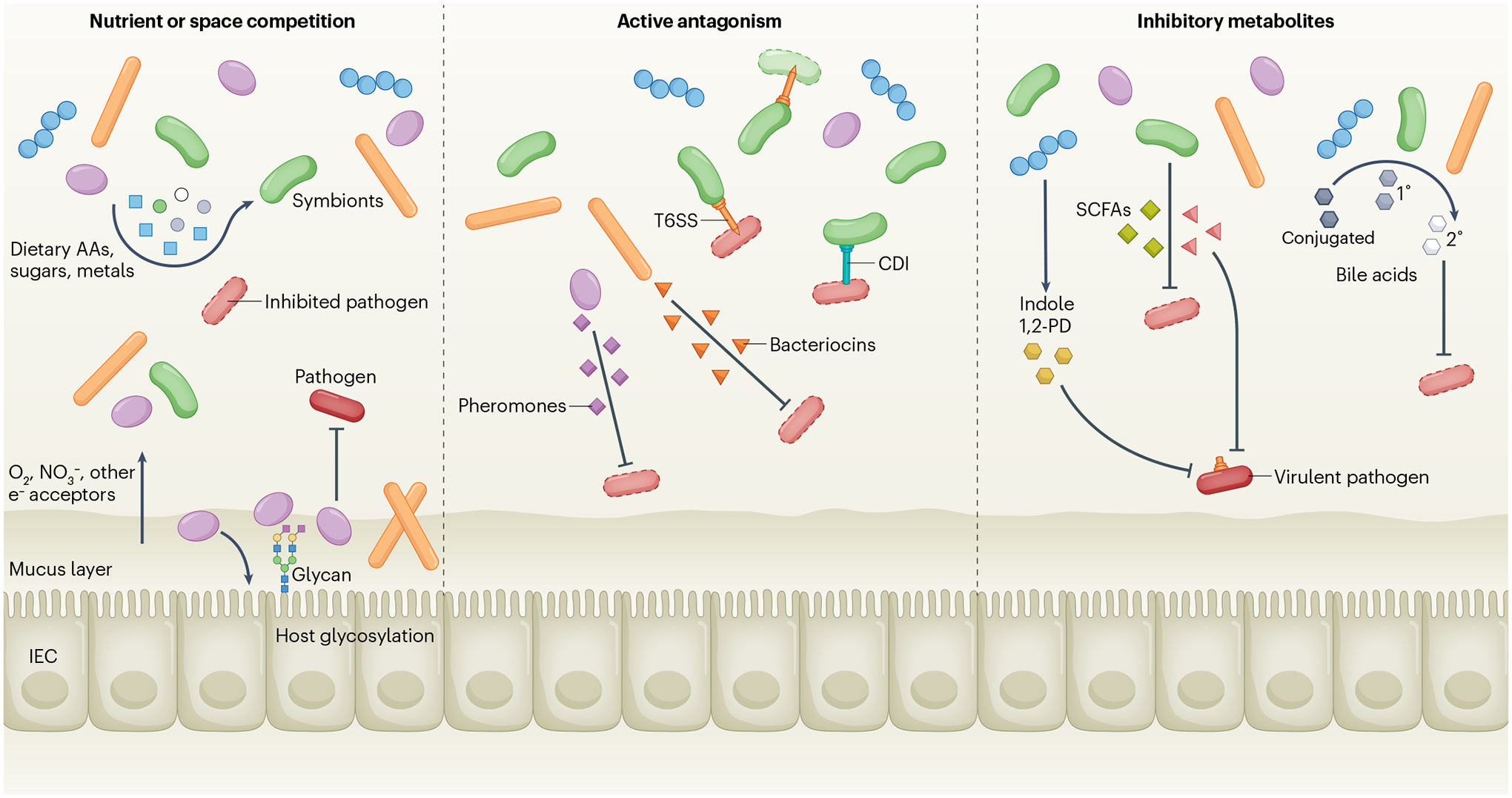
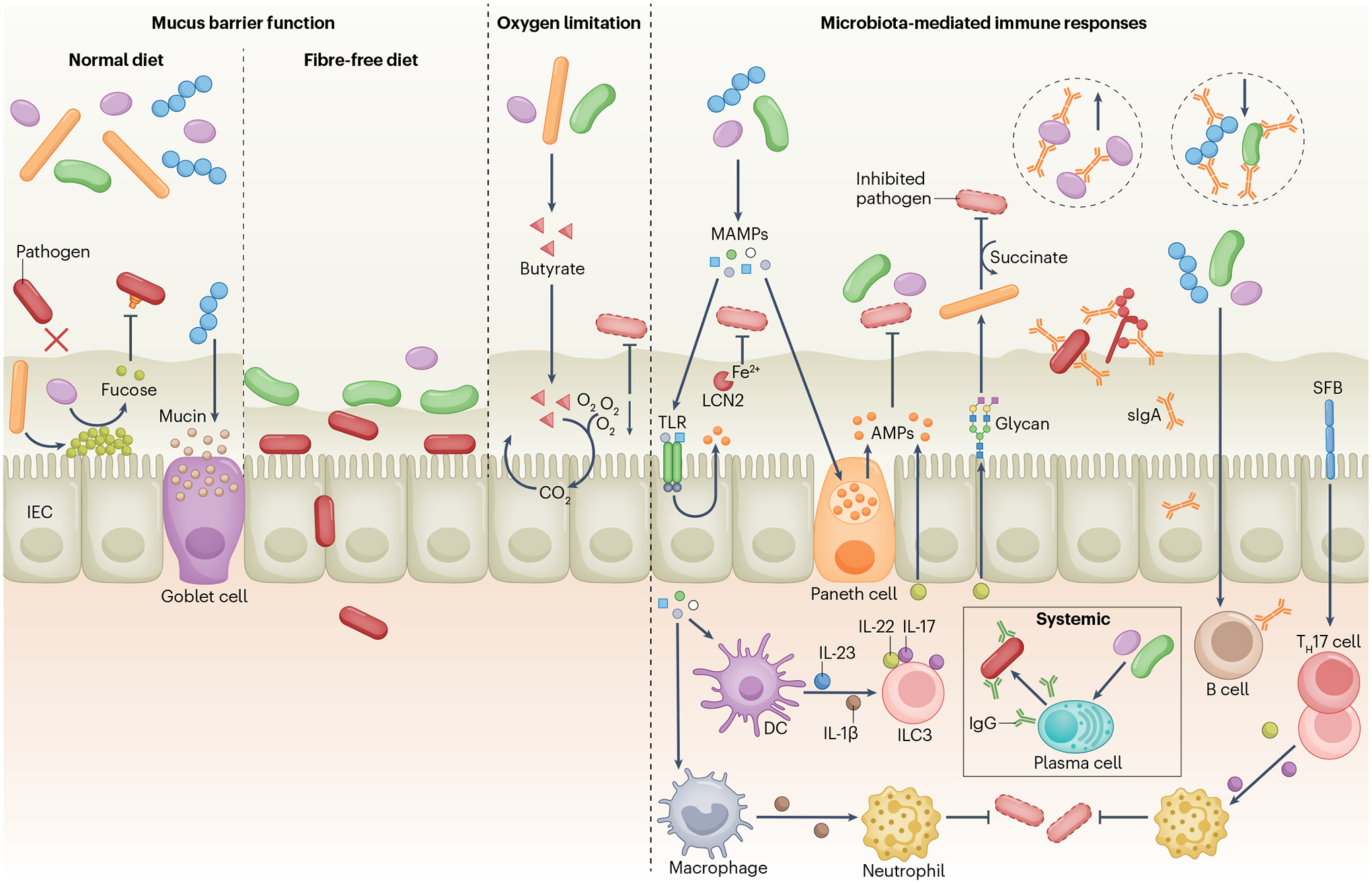
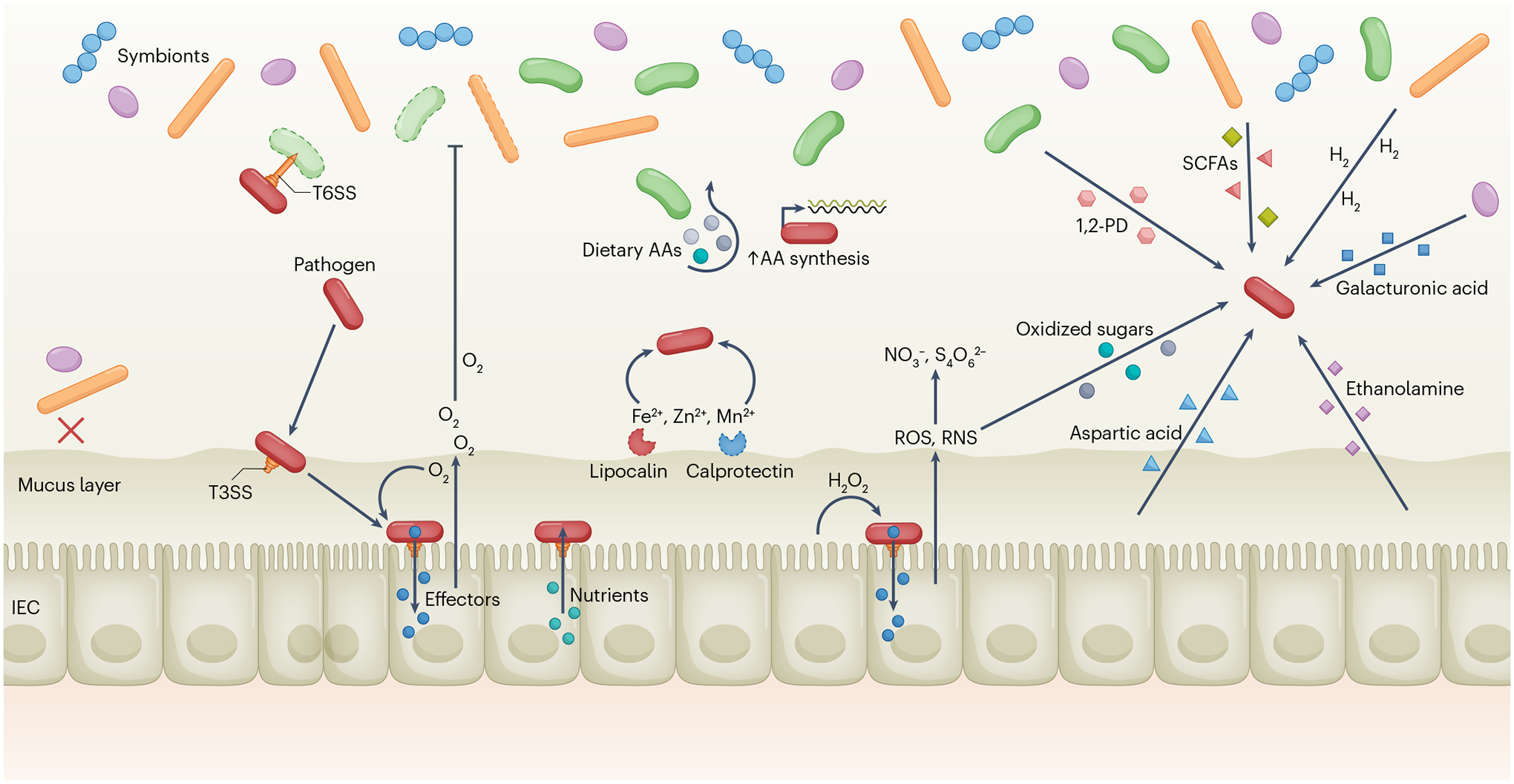
A dense and diverse microbial community inhabits the gut and many epithelial surfaces. Referred to as the microbiota, it co-evolved with the host and is beneficial for many host physiological processes. A major function of these symbiotic microorganisms is protection against pathogen colonization and overgrowth of indigenous pathobionts. Dysbiosis of the normal microbial community increases the risk of pathogen infection and overgrowth of harmful pathobionts. The protective mechanisms conferred by the microbiota are complex and include competitive microbial-microbial interactions and induction of host immune responses. Pathogens, in turn, have evolved multiple strategies to subvert colonization resistance conferred by the microbiota. Understanding the mechanisms by which microbial symbionts limit pathogen colonization should guide the development of new therapeutic approaches to prevent or treat disease.
© 2022. Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
