

Polyglutamine-expanded ATXN7 alters a specific epigenetic signature underlying photoreceptor identity gene expression in SCA7 mouse retinopathy
- PMID: 36539812
- PMCID: PMC9768914
- DOI: 10.1186/s12929-022-00892-1
Polyglutamine-expanded ATXN7 alters a specific epigenetic signature underlying photoreceptor identity gene expression in SCA7 mouse retinopathy
Abstract
Background: Spinocerebellar ataxia type 7 (SCA7) is a neurodegenerative disorder that primarily affects the cerebellum and retina. SCA7 is caused by a polyglutamine expansion in the ATXN7 protein, a subunit of the transcriptional coactivator SAGA that acetylates histone H3 to deposit narrow H3K9ac mark at DNA regulatory elements of active genes. Defective histone acetylation has been presented as a possible cause for gene deregulation in SCA7 mouse models. However, the topography of acetylation defects at the whole genome level and its relationship to changes in gene expression remain to be determined.
Methods: We performed deep RNA-sequencing and chromatin immunoprecipitation coupled to high-throughput sequencing to examine the genome-wide correlation between gene deregulation and alteration of the active transcription marks, e.g. SAGA-related H3K9ac, CBP-related H3K27ac and RNA polymerase II (RNAPII), in a SCA7 mouse retinopathy model.
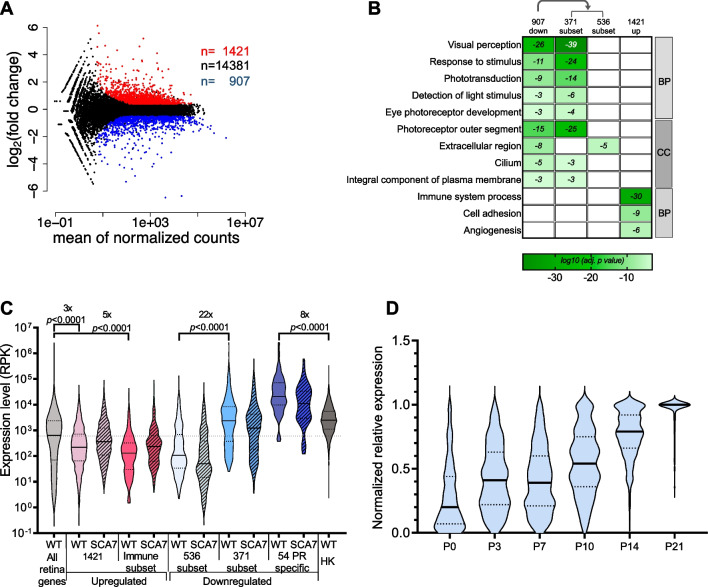
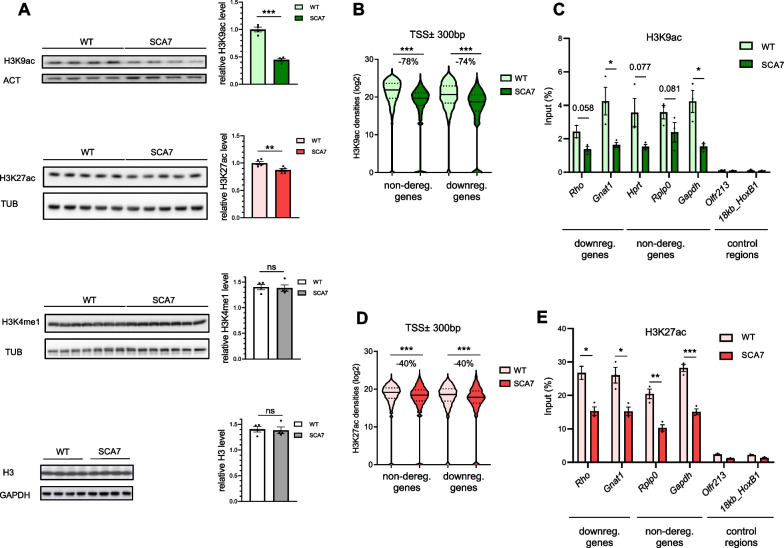
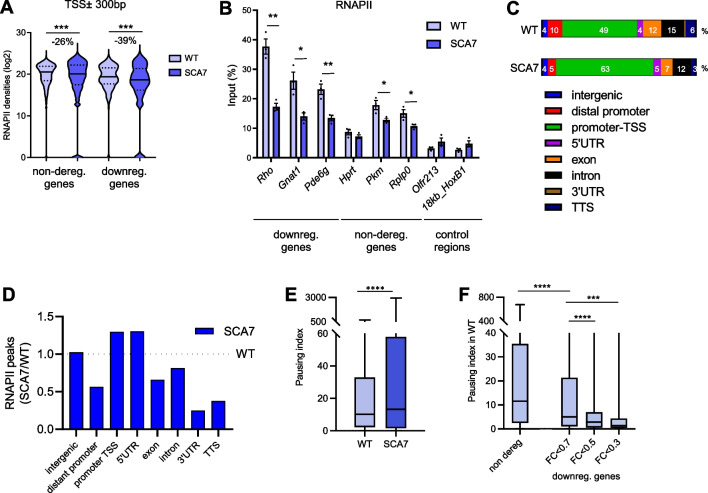
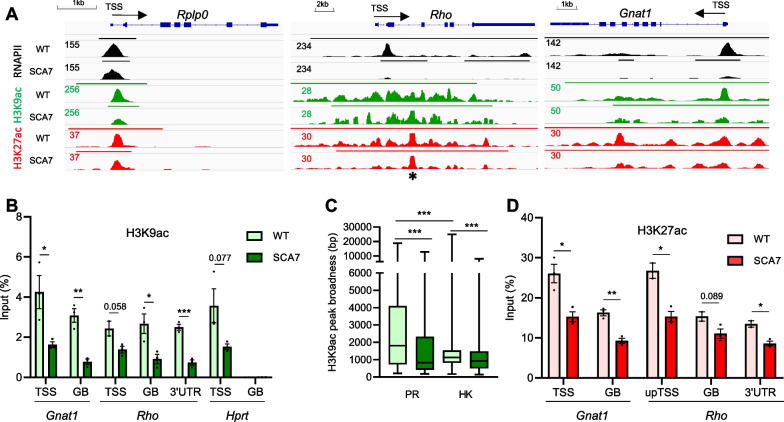
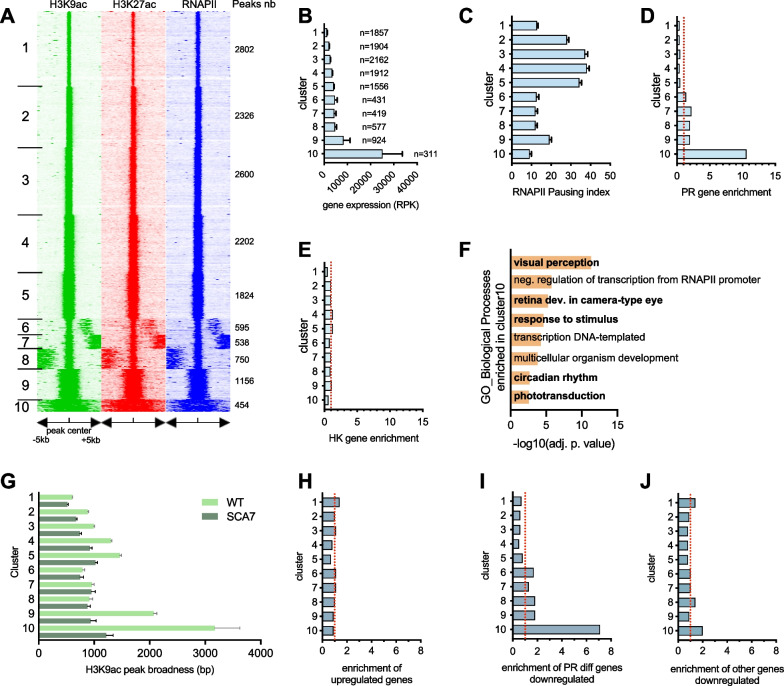
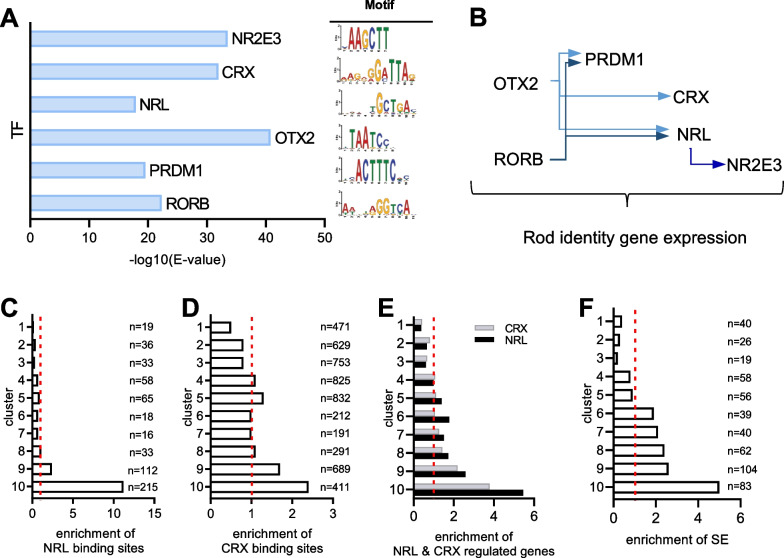
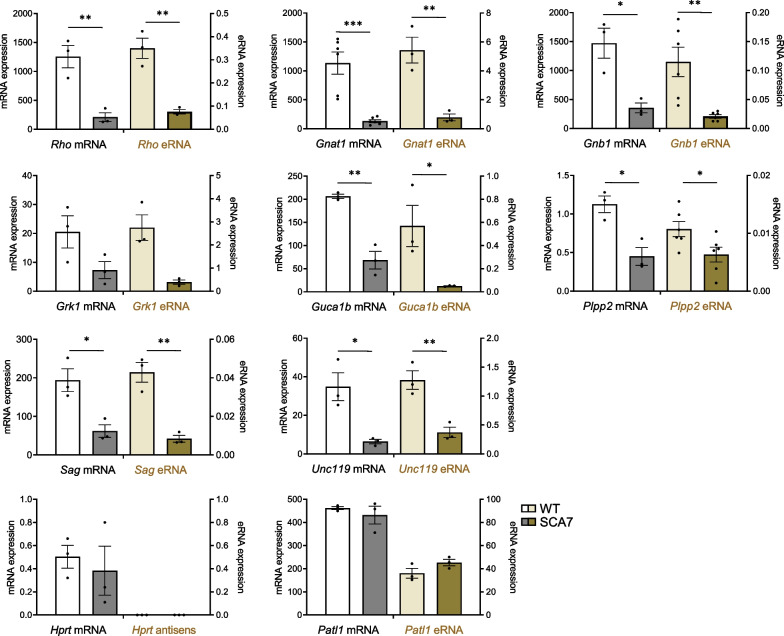
Results: Our analyses revealed that active transcription marks are reduced at most gene promoters in SCA7 retina, while a limited number of genes show changes in expression. We found that SCA7 retinopathy is caused by preferential downregulation of hundreds of highly expressed genes that define morphological and physiological identities of mature photoreceptors. We further uncovered that these photoreceptor genes harbor unusually broad H3K9ac profiles spanning the entire gene bodies and have a low RNAPII pausing. This broad H3K9ac signature co-occurs with other features that delineate superenhancers, including broad H3K27ac, binding sites for photoreceptor specific transcription factors and expression of enhancer-related non-coding RNAs (eRNAs). In SCA7 retina, downregulated photoreceptor genes show decreased H3K9 and H3K27 acetylation and eRNA expression as well as increased RNAPII pausing, suggesting that superenhancer-related features are altered.
Conclusions: Our study thus provides evidence that distinctive epigenetic configurations underlying high expression of cell-type specific genes are preferentially impaired in SCA7, resulting in a defect in the maintenance of identity features of mature photoreceptors. Our results also suggest that continuous SAGA-driven acetylation plays a role in preserving post-mitotic neuronal identity.
Keywords: Epigenomics; Neuronal identity; Photoreceptor dystrophy; SAGA; Spinocerebellar ataxia type 7; Transcriptomics.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Karmodiya K, Krebs AR, Oulad-Abdelghani M, Kimura H, Tora L. H3K9 and H3K14 acetylation co-occur at many gene regulatory elements, while H3K14ac marks a subset of inactive inducible promoters in mouse embryonic stem cells. BMC Genomics. 2012;13:424. doi: 10.1186/1471-2164-13-424. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
