

Alternative splicing liberates a cryptic cytoplasmic isoform of mitochondrial MECR that antagonizes influenza virus
- PMID: 36542656
- PMCID: PMC9815647
- DOI: 10.1371/journal.pbio.3001934
Alternative splicing liberates a cryptic cytoplasmic isoform of mitochondrial MECR that antagonizes influenza virus
Abstract
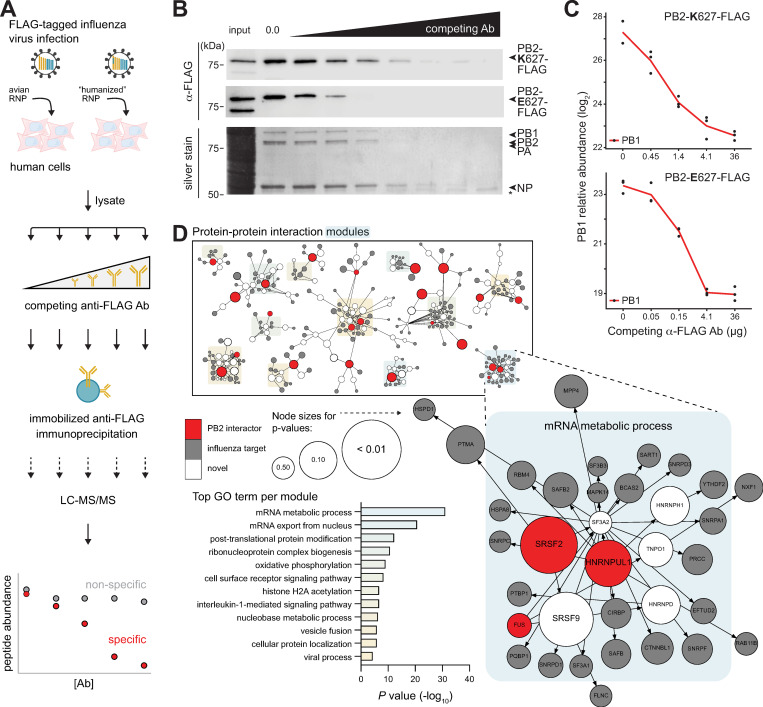
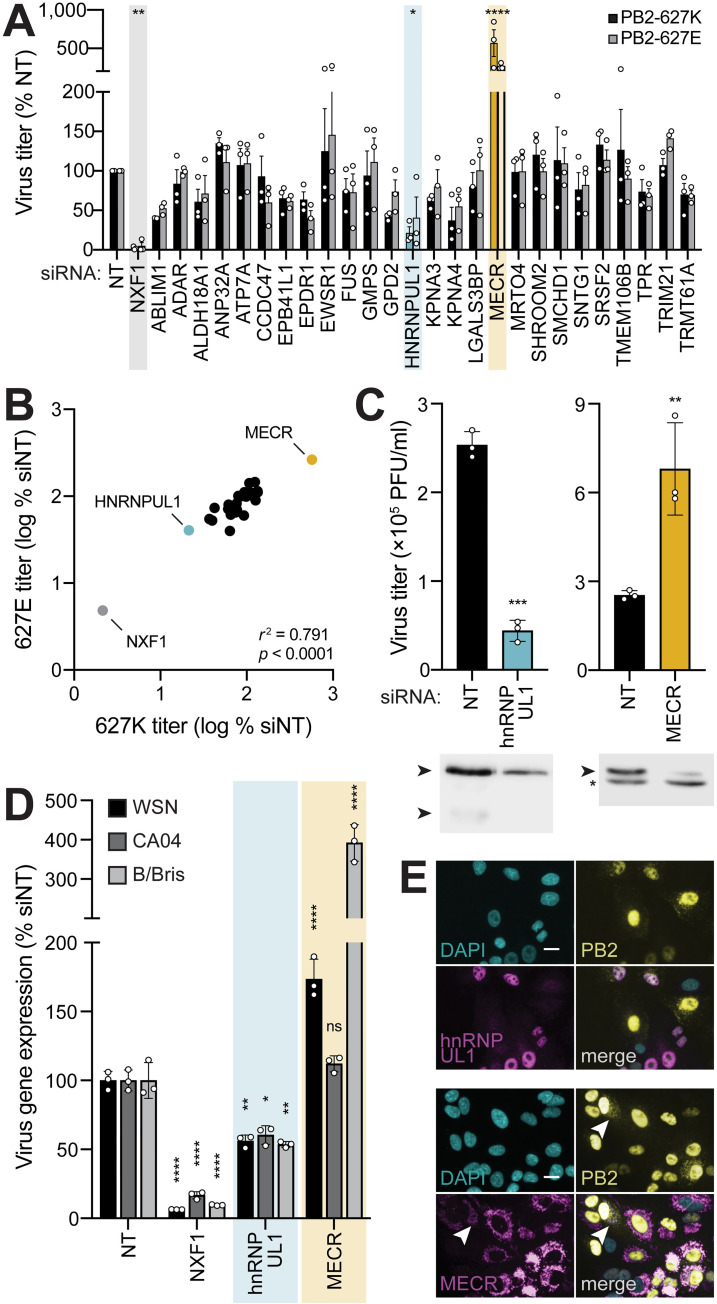
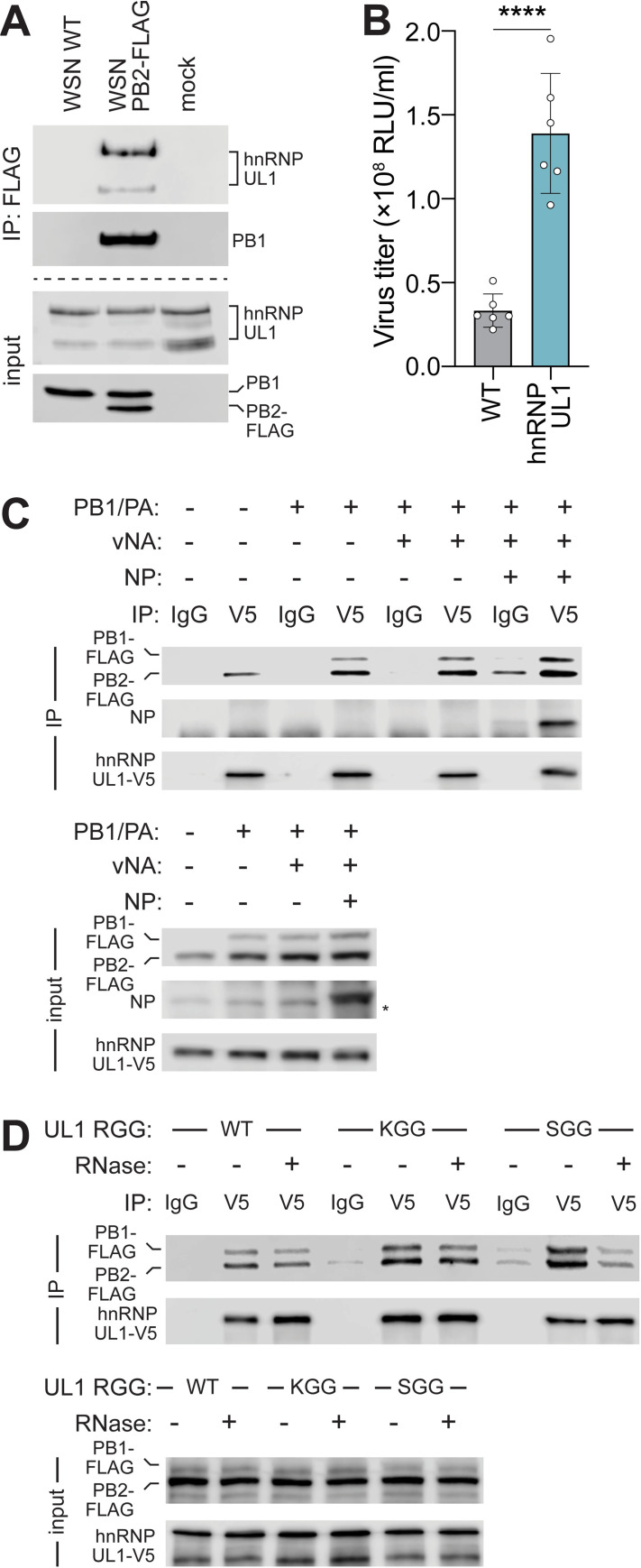
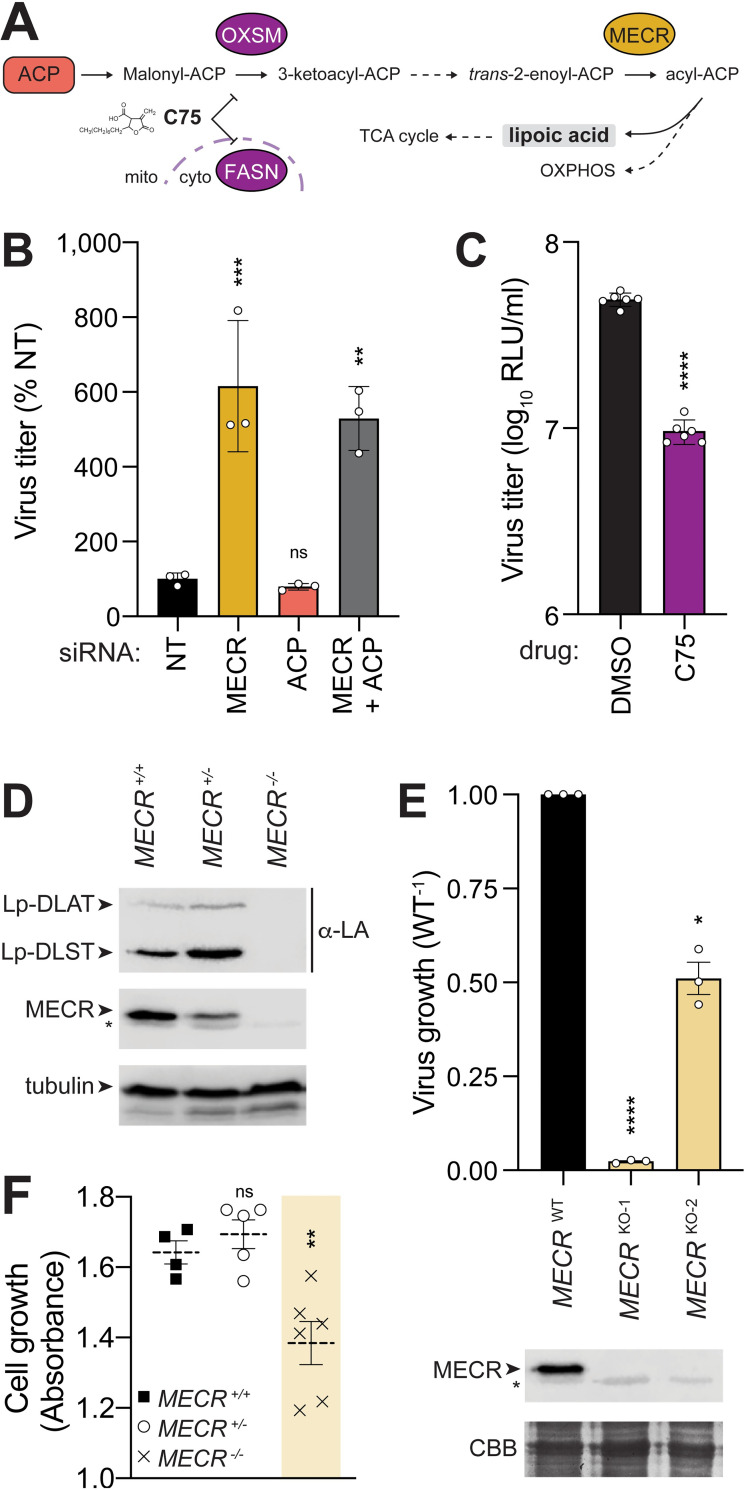
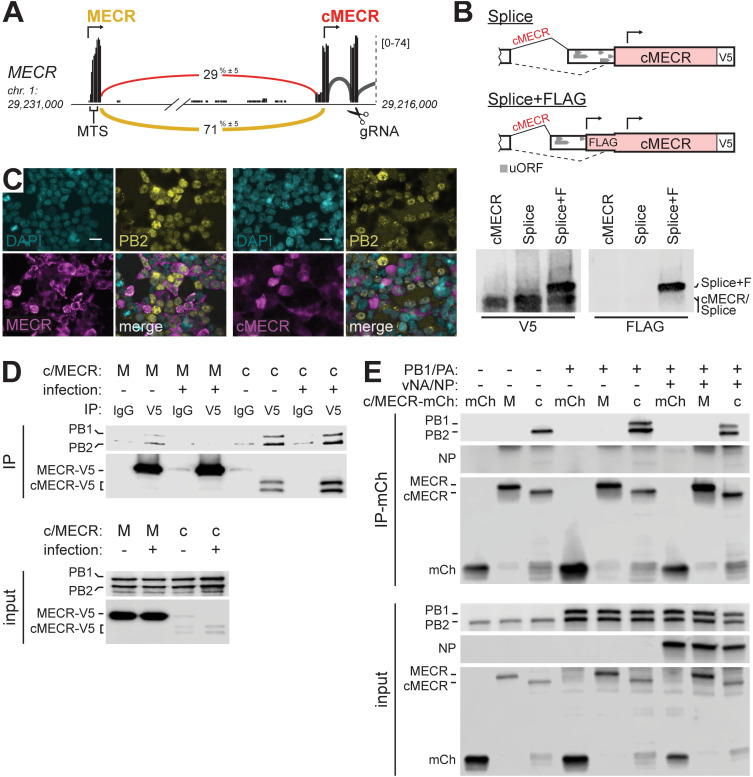
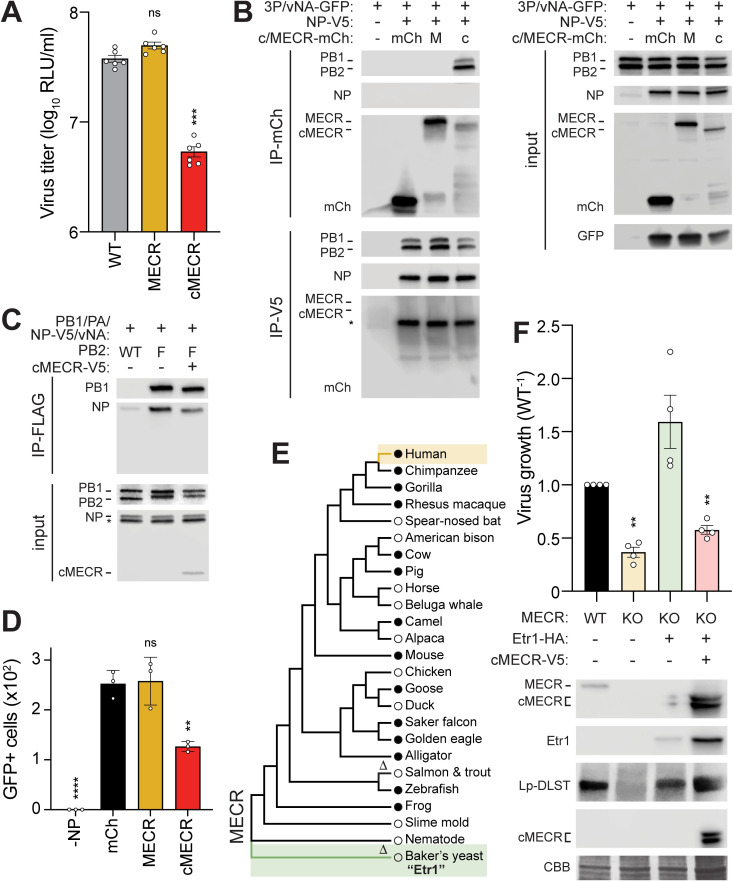
Viruses must balance their reliance on host cell machinery for replication while avoiding host defense. Influenza A viruses are zoonotic agents that frequently switch hosts, causing localized outbreaks with the potential for larger pandemics. The host range of influenza virus is limited by the need for successful interactions between the virus and cellular partners. Here we used immunocompetitive capture-mass spectrometry to identify cellular proteins that interact with human- and avian-style viral polymerases. We focused on the proviral activity of heterogenous nuclear ribonuclear protein U-like 1 (hnRNP UL1) and the antiviral activity of mitochondrial enoyl CoA-reductase (MECR). MECR is localized to mitochondria where it functions in mitochondrial fatty acid synthesis (mtFAS). While a small fraction of the polymerase subunit PB2 localizes to the mitochondria, PB2 did not interact with full-length MECR. By contrast, a minor splice variant produces cytoplasmic MECR (cMECR). Ectopic expression of cMECR shows that it binds the viral polymerase and suppresses viral replication by blocking assembly of viral ribonucleoprotein complexes (RNPs). MECR ablation through genome editing or drug treatment is detrimental for cell health, creating a generic block to virus replication. Using the yeast homolog Etr1 to supply the metabolic functions of MECR in MECR-null cells, we showed that specific antiviral activity is independent of mtFAS and is reconstituted by expressing cMECR. Thus, we propose a strategy where alternative splicing produces a cryptic antiviral protein that is embedded within a key metabolic enzyme.
Copyright: © 2022 Baker et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: AM is an editorial board member for PLoS Biology and PLoS Pathogens. HM, MT, SG, JSP, AA and HJ were employees of F. Hoffmann-La Roche when performing this work. No other authors declare a competing interest.
Figures
References
-
- Garten R, Blanton L, Elal AIA, Alabi N, Barnes J, Biggerstaff M, et al.. Update: Influenza Activity in the United States During the 2017–18 Season and Composition of the 2018–19 Influenza Vaccine. MMWR Morbidity and Mortality Weekly Report. 2018;67:634–642. doi: 10.15585/mmwr.mm6722a4 - DOI - PMC - PubMed
-
- Van Valen L. The Red Queen lives. Nature. 1976;260:575. doi: 10.1038/260575a0 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
