

A cortico-collicular circuit for orienting to shelter during escape
- PMID: 36544025
- PMCID: PMC7614651
- DOI: 10.1038/s41586-022-05553-9
A cortico-collicular circuit for orienting to shelter during escape
Abstract
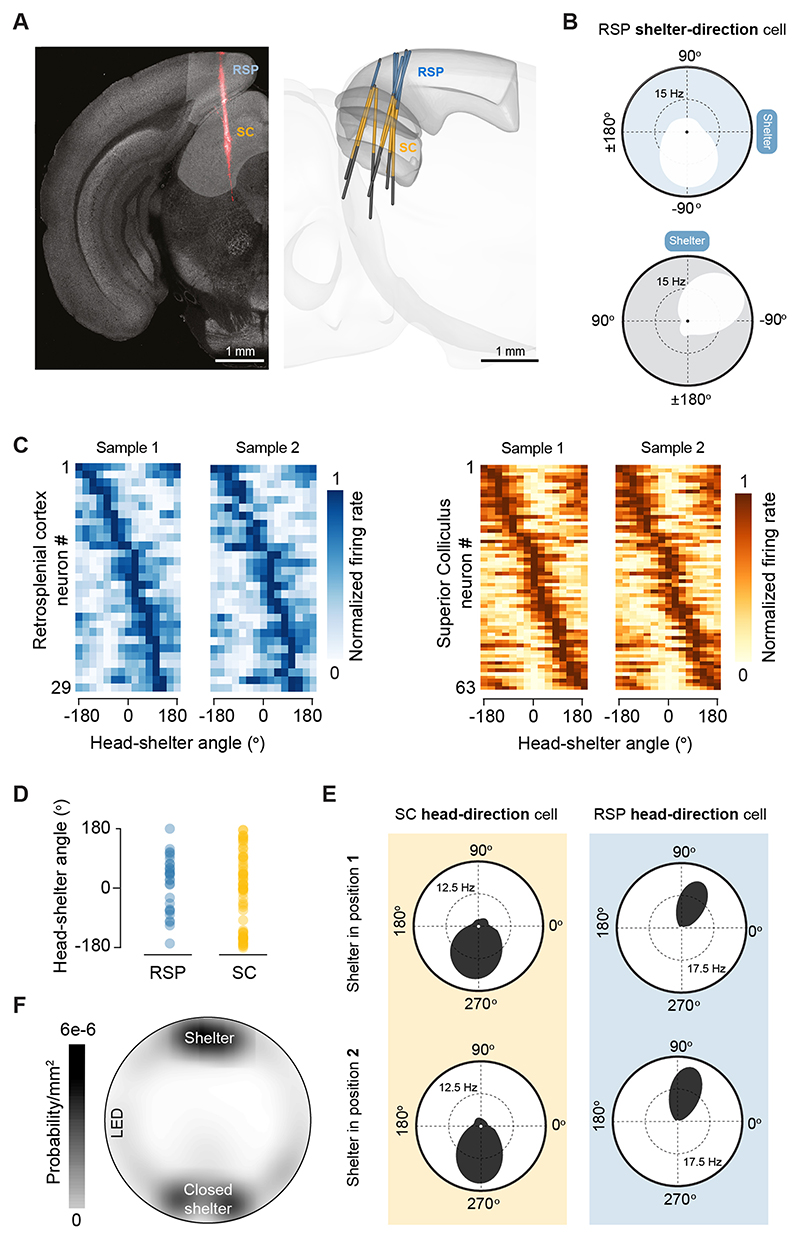
When faced with predatory threats, escape towards shelter is an adaptive action that offers long-term protection against the attacker. Animals rely on knowledge of safe locations in the environment to instinctively execute rapid shelter-directed escape actions1,2. Although previous work has identified neural mechanisms of escape initiation3,4, it is not known how the escape circuit incorporates spatial information to execute rapid flights along the most efficient route to shelter. Here we show that the mouse retrosplenial cortex (RSP) and superior colliculus (SC) form a circuit that encodes the shelter-direction vector and is specifically required for accurately orienting to shelter during escape. Shelter direction is encoded in RSP and SC neurons in egocentric coordinates and SC shelter-direction tuning depends on RSP activity. Inactivation of the RSP-SC pathway disrupts the orientation to shelter and causes escapes away from the optimal shelter-directed route, but does not lead to generic deficits in orientation or spatial navigation. We find that the RSP and SC are monosynaptically connected and form a feedforward lateral inhibition microcircuit that strongly drives the inhibitory collicular network because of higher RSP input convergence and synaptic integration efficiency in inhibitory SC neurons. This results in broad shelter-direction tuning in inhibitory SC neurons and sharply tuned excitatory SC neurons. These findings are recapitulated by a biologically constrained spiking network model in which RSP input to the local SC recurrent ring architecture generates a circular shelter-direction map. We propose that this RSP-SC circuit might be specialized for generating collicular representations of memorized spatial goals that are readily accessible to the motor system during escape, or more broadly, during navigation when the goal must be reached as fast as possible.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures


















References
-
- van der Meer MAA, Richmond Z, Braga RM, Wood ER, Dudchenko PA. Evidence for the Use of an Internal Sense of Direction in Homing. Behavioral Neuroscience. 2010;124:164–169. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
