

Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination
- PMID: 36548397
- PMCID: PMC9847566
- DOI: 10.1126/sciimmunol.ade2798
Class switch toward noninflammatory, spike-specific IgG4 antibodies after repeated SARS-CoV-2 mRNA vaccination
Abstract
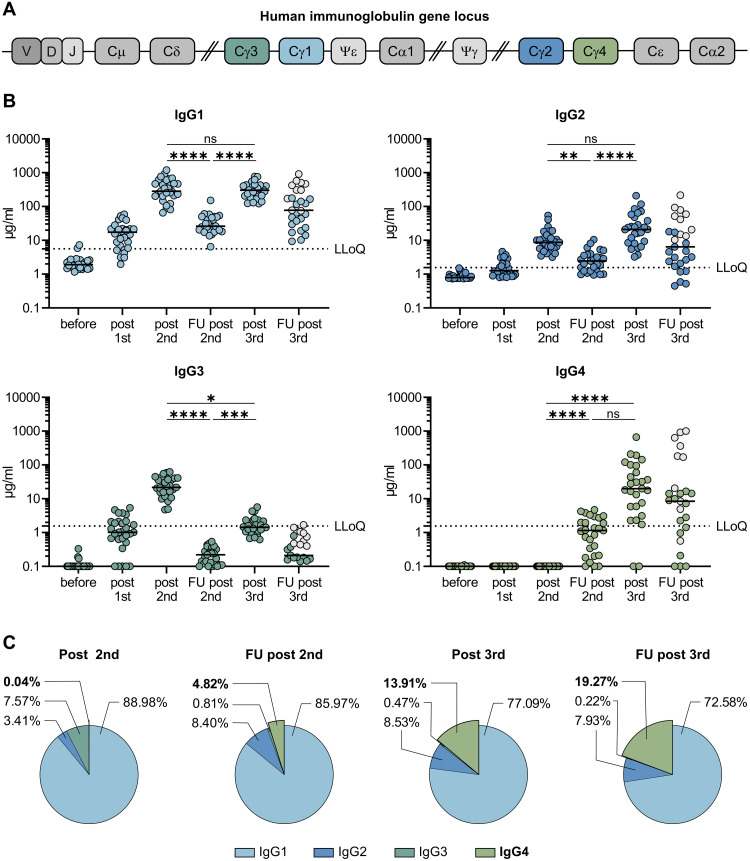
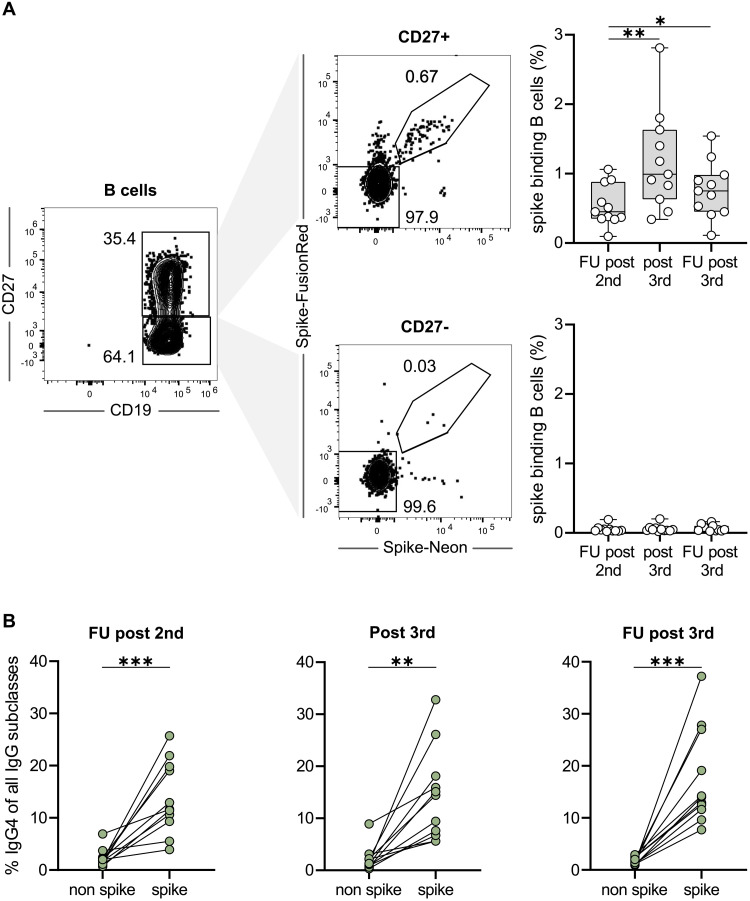
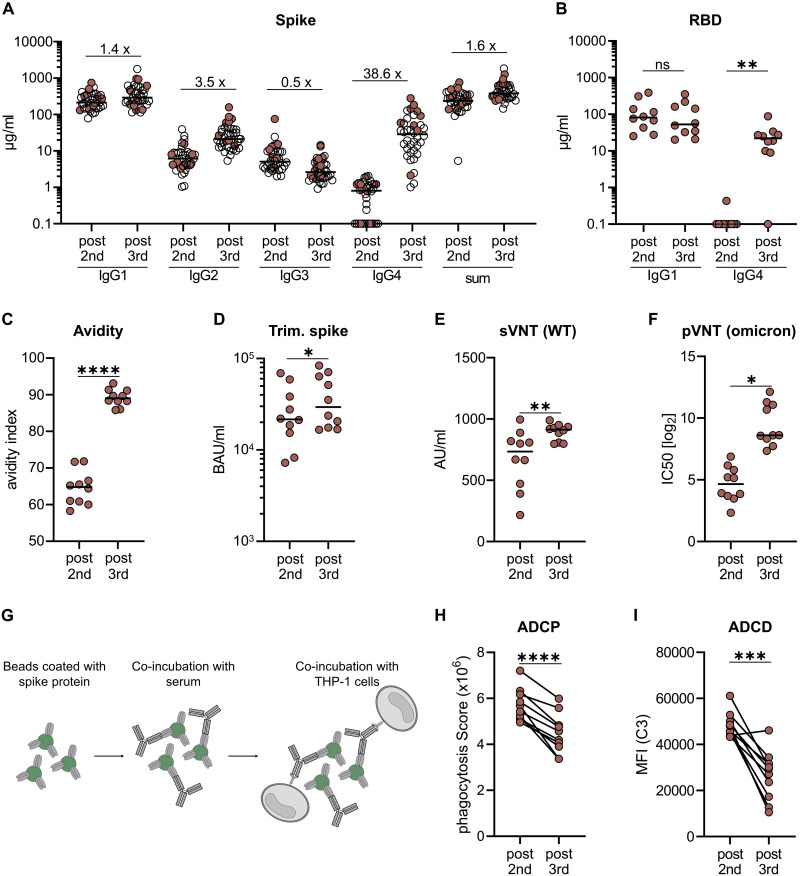
RNA vaccines are efficient preventive measures to combat the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) pandemic. High levels of neutralizing SARS-CoV-2 antibodies are an important component of vaccine-induced immunity. Shortly after the initial two mRNA vaccine doses, the immunoglobulin G (IgG) response mainly consists of the proinflammatory subclasses IgG1 and IgG3. Here, we report that several months after the second vaccination, SARS-CoV-2-specific antibodies were increasingly composed of noninflammatory IgG4, which were further boosted by a third mRNA vaccination and/or SARS-CoV-2 variant breakthrough infections. IgG4 antibodies among all spike-specific IgG antibodies rose, on average, from 0.04% shortly after the second vaccination to 19.27% late after the third vaccination. This induction of IgG4 antibodies was not observed after homologous or heterologous SARS-CoV-2 vaccination with adenoviral vectors. Single-cell sequencing and flow cytometry revealed substantial frequencies of IgG4-switched B cells within the spike-binding memory B cell population [median of 14.4%; interquartile range (IQR) of 6.7 to 18.1%] compared with the overall memory B cell repertoire (median of 1.3%; IQR of 0.9 to 2.2%) after three immunizations. This class switch was associated with a reduced capacity of the spike-specific antibodies to mediate antibody-dependent cellular phagocytosis and complement deposition. Because Fc-mediated effector functions are critical for antiviral immunity, these findings may have consequences for the choice and timing of vaccination regimens using mRNA vaccines, including future booster immunizations against SARS-CoV-2.
Figures
Comment in
-
Is it bad, is it good, or is IgG4 just misunderstood?Sci Immunol. 2023 Mar 31;8(81):eadg7327. doi: 10.1126/sciimmunol.adg7327. Epub 2023 Mar 24. Sci Immunol. 2023. PMID: 36749191
References
-
- Meslé M. M., Brown J., Mook P., Hagan J., Pastore R., Bundle N., Spiteri G., Ravasi G., Nicolay N., Andrews N., Dykhanovska T., Mossong J., Sadkowska-Todys M., Nikiforova R., Riccardo F., Meijerink H., Mazagatos C., Kyncl J., McMenamin J., Melillo T., Kaoustou S., Lévy-Bruhl D., Haarhuis F., Rich R., Kall M., Nitzan D., Smallwood C., Pebody R. G., Estimated number of deaths directly averted in people 60 years and older as a result of COVID-19 vaccination in the WHO European Region, December 2020 to November 2021. Euro. Surveill. 26, 2101021 (2021). - PMC - PubMed
-
- Polack F. P., Thomas S. J., Kitchin N., Absalon J., Gurtman A., Lockhart S., Perez J. L., Pérez Marc G., Moreira E. D., Zerbini C., Bailey R., Swanson K. A., Roychoudhury S., Koury K., Li P., Kalina W. V., Cooper D., Frenck R. W., Hammitt L. L., Türeci Ö., Nell H., Schaefer A., Ünal S., Tresnan D. B., Mather S., Dormitzer P. R., Şahin U., Jansen K. U., Gruber W. C.; C4591001 Clinical Trial Group , Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 383, 2603–2615 (2020). - PMC - PubMed
-
- Baden L. R., El Sahly H. M., Essink B., Kotloff K., Frey S., Novak R., Diemert D., Spector S. A., Rouphael N., Creech C. B., McGettigan J., Khetan S., Segall N., Solis J., Brosz A., Fierro C., Schwartz H., Neuzil K., Corey L., Gilbert P., Janes H., Follmann D., Marovich M., Mascola J., Polakowski L., Ledgerwood J., Graham B. S., Bennett H., Pajon R., Knightly C., Leav B., Deng W., Zhou H., Han S., Ivarsson M., Miller J., Zaks T.; COVE Study Group , Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 384, 403–416 (2021). - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
