

Benchmarking transcriptional host response signatures for infection diagnosis
- PMID: 36549274
- PMCID: PMC9768893
- DOI: 10.1016/j.cels.2022.11.007
Benchmarking transcriptional host response signatures for infection diagnosis
Abstract
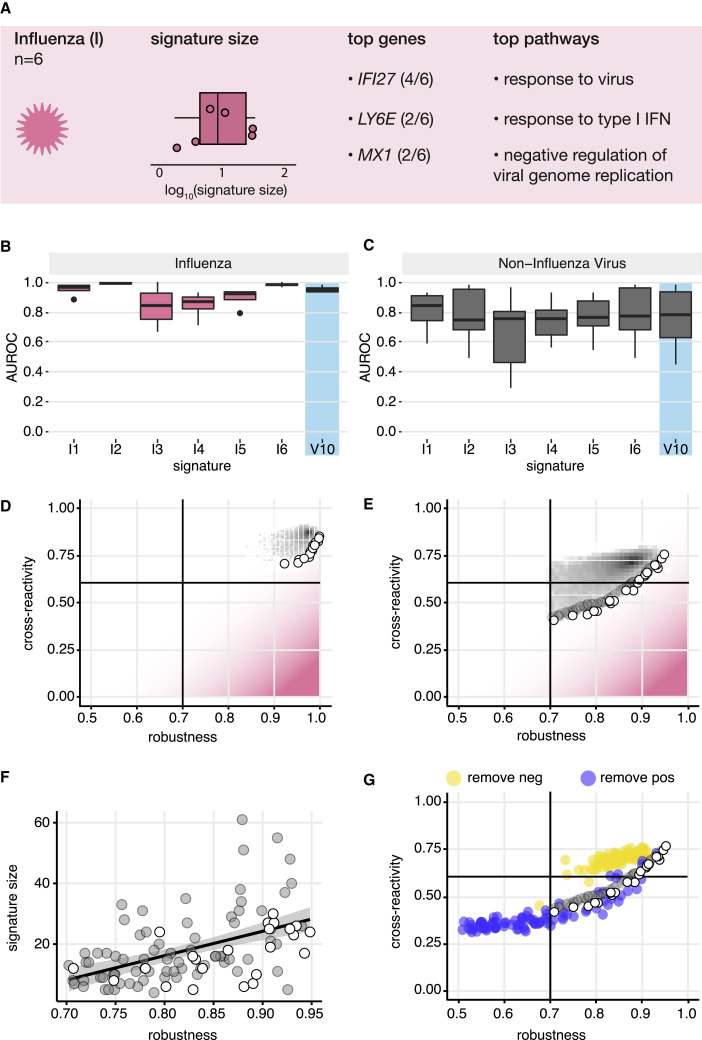
Identification of host transcriptional response signatures has emerged as a new paradigm for infection diagnosis. For clinical applications, signatures must robustly detect the pathogen of interest without cross-reacting with unintended conditions. To evaluate the performance of infectious disease signatures, we developed a framework that includes a compendium of 17,105 transcriptional profiles capturing infectious and non-infectious conditions and a standardized methodology to assess robustness and cross-reactivity. Applied to 30 published signatures of infection, the analysis showed that signatures were generally robust in detecting viral and bacterial infections in independent data. Asymptomatic and chronic infections were also detectable, albeit with decreased performance. However, many signatures were cross-reactive with unintended infections and aging. In general, we found robustness and cross-reactivity to be conflicting objectives, and we identified signature properties associated with this trade-off. The data compendium and evaluation framework developed here provide a foundation for the development of signatures for clinical application. A record of this paper's transparent peer review process is included in the supplemental information.
Keywords: aging; bacteria; cross-reactivity; data compendium; infection diagnosis; influenza signature; non-infectious conditions; robustness; signature evaluation framework; transcriptional host response signature; virus.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Icahn School of Medicine at Mount Sinai has submitted a provisional patent related to this work. A.C., D.G.C., S.C.S., S.H.K., and E.Z. are inventors of the technology filed through ISMMS related to this manuscript. S.H.K. receives consulting fees from Peraton.
Figures






Comment on
-
Multi-objective optimization identifies a specific and interpretable COVID-19 host response signature.Cell Syst. 2022 Dec 21;13(12):989-1001.e8. doi: 10.1016/j.cels.2022.11.008. Cell Syst. 2022. PMID: 36549275
References
-
- Ferrer R., Martin-Loeches I., Phillips G., Osborn T.M., Townsend S., Dellinger R.P., Artigas A., Schorr C., Levy M.M. Empiric antibiotic treatment reduces mortality in severe sepsis and septic shock from the first hour: results from a guideline-based performance improvement program. Crit. Care Med. 2014;42:1749–1755. doi: 10.1097/CCM.0000000000000330. - DOI - PubMed
-
- CDC . Centers for Disease Control and Prevention; 2020. Antibiotic Resistance is a National Priority.https://www.cdc.gov/drugresistance/us-activities.html
-
- Self W.H., Balk R.A., Grijalva C.G., Williams D.J., Zhu Y., Anderson E.J., Waterer G.W., Courtney D.M., Bramley A.M., Trabue C., et al. Procalcitonin as a marker of etiology in adults hospitalized with community-acquired pneumonia. Clin. Infect. Dis. 2017;65:183–190. doi: 10.1093/cid/cix317. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
