

Structural insights into the covalent regulation of PAPP-A activity by proMBP and STC2
- PMID: 36550107
- PMCID: PMC9780223
- DOI: 10.1038/s41421-022-00502-2
Structural insights into the covalent regulation of PAPP-A activity by proMBP and STC2
Abstract
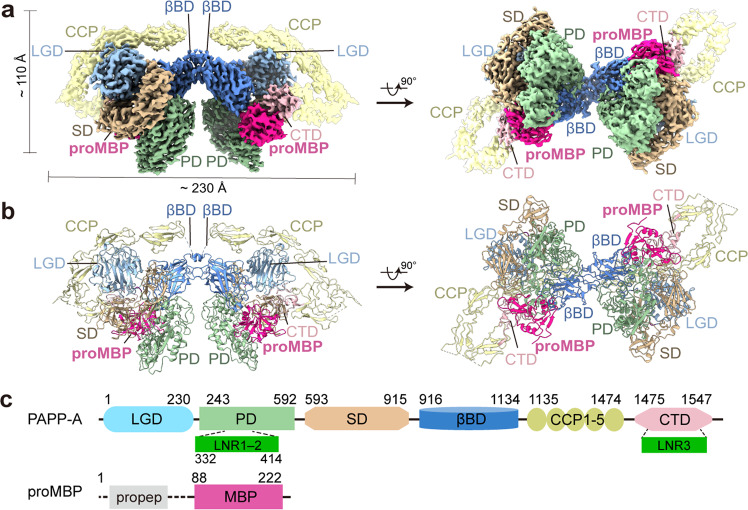
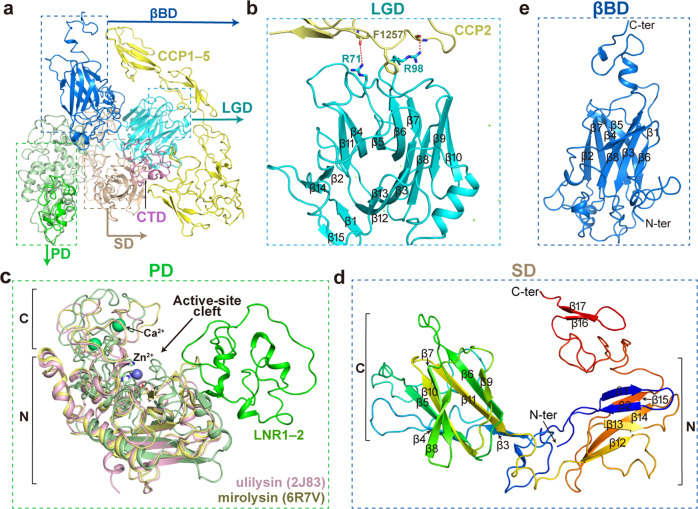
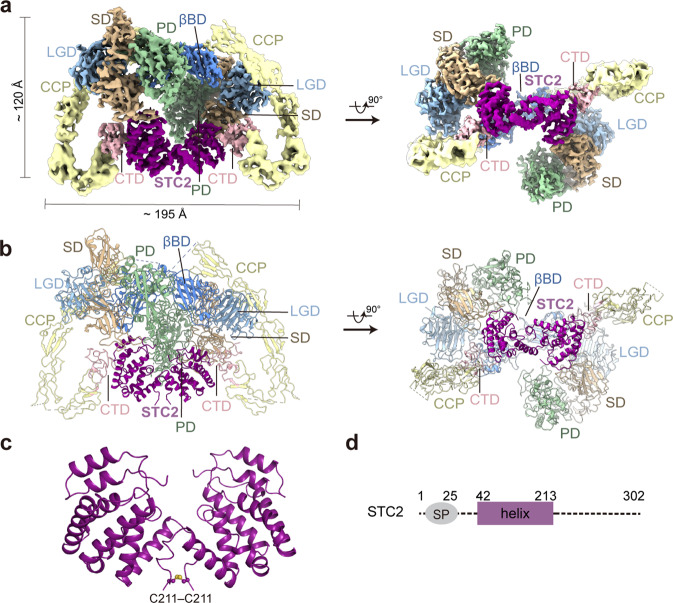
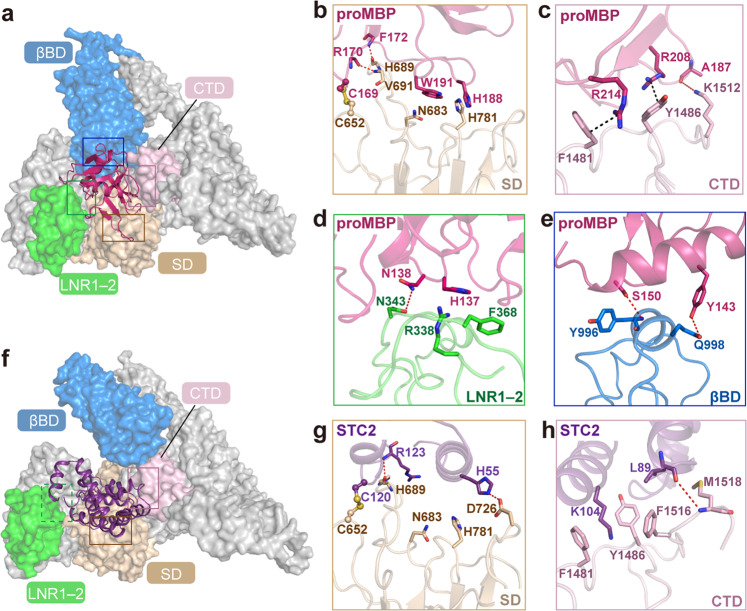
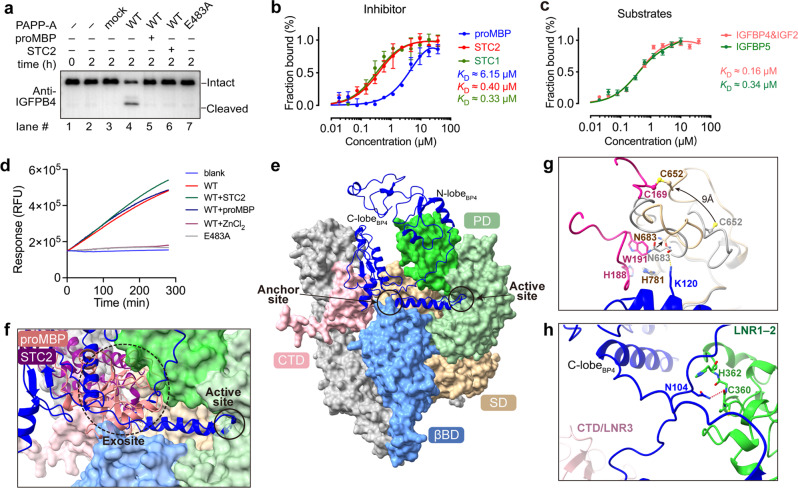
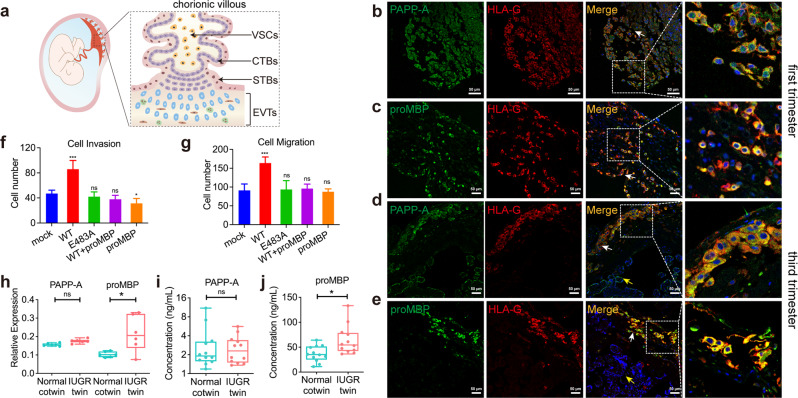
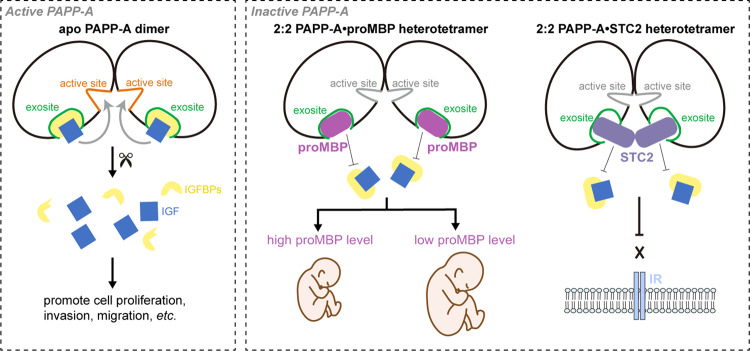
Originally discovered in the circulation of pregnant women as a protein secreted by placental trophoblasts, the metalloprotease pregnancy-associated plasma protein A (PAPP-A) is also widely expressed by many other tissues. It cleaves insulin-like growth factor-binding proteins (IGFBPs) to increase the bioavailability of IGFs and plays essential roles in multiple growth-promoting processes. While the vast majority of the circulatory PAPP-A in pregnancy is proteolytically inactive due to covalent inhibition by proform of eosinophil major basic protein (proMBP), the activity of PAPP-A can also be covalently inhibited by another less characterized modulator, stanniocalcin-2 (STC2). However, the structural basis of PAPP-A proteolysis and the mechanistic differences between these two modulators are poorly understood. Here we present two cryo-EM structures of endogenous purified PAPP-A in complex with either proMBP or STC2. Both modulators form 2:2 heterotetramer with PAPP-A and establish extensive interactions with multiple domains of PAPP-A that are distal to the catalytic cleft. This exosite-binding property results in a steric hindrance to prevent the binding and cleavage of IGFBPs, while the IGFBP linker region-derived peptides harboring the cleavage sites are no longer sensitive to the modulator treatment. Functional investigation into proMBP-mediated PAPP-A regulation in selective intrauterine growth restriction (sIUGR) pregnancy elucidates that PAPP-A and proMBP collaboratively regulate extravillous trophoblast invasion and the consequent fetal growth. Collectively, our work reveals a novel covalent exosite-competitive inhibition mechanism of PAPP-A and its regulatory effect on placental function.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Identification and regulation of the IGFBP-4 protease and its physiological inhibitor in human trophoblasts and endometrial stroma: evidence for paracrine regulation of IGF-II bioavailability in the placental bed during human implantation.J Clin Endocrinol Metab. 2002 May;87(5):2359-66. doi: 10.1210/jcem.87.5.8448. J Clin Endocrinol Metab. 2002. PMID: 11994388
-
Inhibition of proteolysis by the proform of eosinophil major basic protein (proMBP) requires covalent binding to its target proteinase.FEBS Lett. 2004 Feb 27;560(1-3):147-52. doi: 10.1016/S0014-5793(04)00095-X. FEBS Lett. 2004. PMID: 14988014
-
Expression of recombinant human pregnancy-associated plasma protein-A and identification of the proform of eosinophil major basic protein as its physiological inhibitor.J Biol Chem. 2000 Oct 6;275(40):31128-33. doi: 10.1074/jbc.M001384200. J Biol Chem. 2000. PMID: 10913121
-
PAPP-A and the IGF system.Ann Endocrinol (Paris). 2016 Jun;77(2):90-6. doi: 10.1016/j.ando.2016.04.015. Epub 2016 May 5. Ann Endocrinol (Paris). 2016. PMID: 27155776 Review.
-
Pregnancy-associated plasma proteins and Stanniocalcin-2 - Novel players controlling IGF-I physiology.Growth Horm IGF Res. 2020 Aug-Oct;53-54:101330. doi: 10.1016/j.ghir.2020.101330. Epub 2020 Jul 4. Growth Horm IGF Res. 2020. PMID: 32693362 Review.
Cited by
-
In situ crystalline structure of the human eosinophil major basic protein-1.bioRxiv [Preprint]. 2024 Oct 11:2024.10.09.617336. doi: 10.1101/2024.10.09.617336. bioRxiv. 2024. PMID: 39416224 Free PMC article. Preprint.
-
Unveiling the Role of Pregnancy-Associated Plasma Protein A (PAPP-A) in Pregnancy-Associated Breast Cancer: A Comprehensive Review.Cureus. 2024 Mar 16;16(3):e56269. doi: 10.7759/cureus.56269. eCollection 2024 Mar. Cureus. 2024. PMID: 38623138 Free PMC article. Review.
-
Cryo-EM structure of human PAPP-A2 and mechanism of substrate recognition.Commun Chem. 2023 Oct 28;6(1):234. doi: 10.1038/s42004-023-01032-y. Commun Chem. 2023. PMID: 37898658 Free PMC article.
-
Stanniocalcin 2 governs cancer cell adaptation to nutrient insufficiency through alleviation of oxidative stress.Res Sq [Preprint]. 2024 Feb 27:rs.3.rs-3904465. doi: 10.21203/rs.3.rs-3904465/v1. Res Sq. 2024. Update in: Cell Death Dis. 2024 Aug 6;15(8):567. doi: 10.1038/s41419-024-06961-7. PMID: 38464261 Free PMC article. Updated. Preprint.
-
Regulation and function of insulin and insulin-like growth factor receptor signalling.Nat Rev Mol Cell Biol. 2025 Jul;26(7):558-580. doi: 10.1038/s41580-025-00826-3. Epub 2025 Feb 10. Nat Rev Mol Cell Biol. 2025. PMID: 39930003 Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
