

The Role of Membrane Affinity and Binding Modes in Alpha-Synuclein Regulation of Vesicle Release and Trafficking
- PMID: 36551244
- PMCID: PMC9775087
- DOI: 10.3390/biom12121816
The Role of Membrane Affinity and Binding Modes in Alpha-Synuclein Regulation of Vesicle Release and Trafficking
Abstract
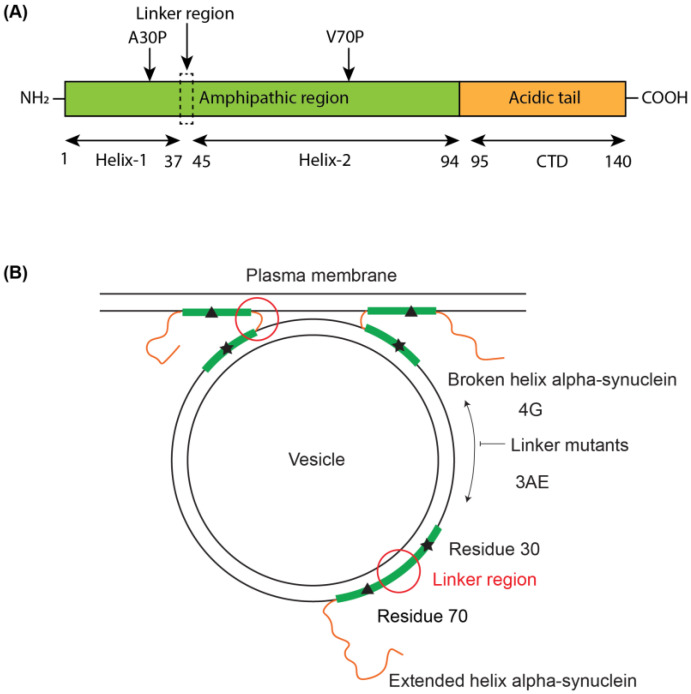
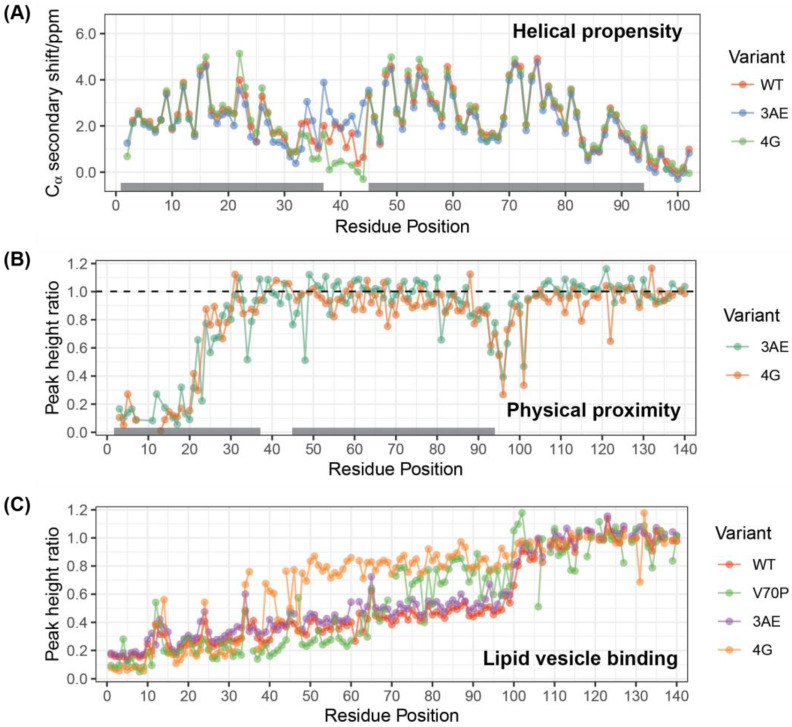
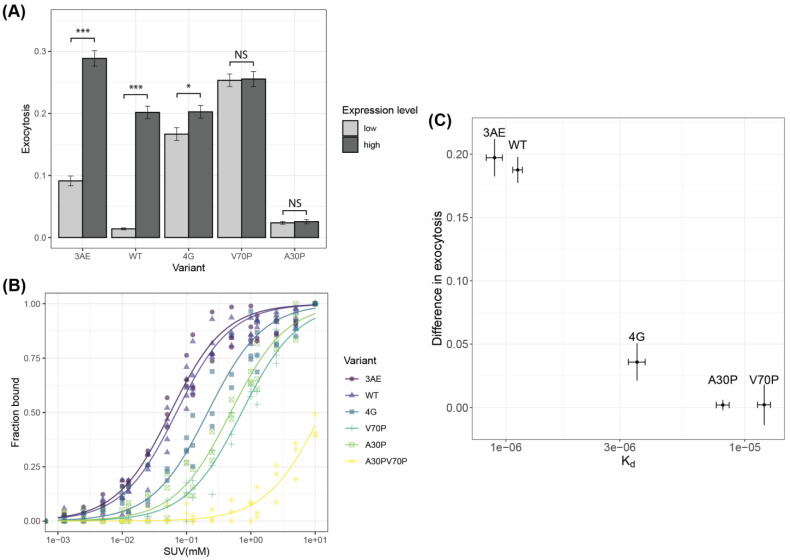
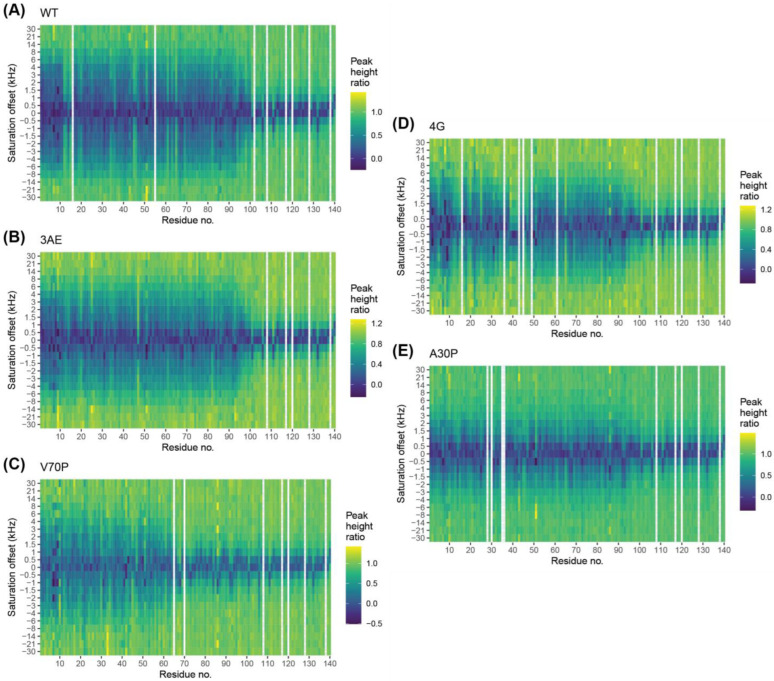
Alpha-synuclein is a presynaptic protein linked to Parkinson's disease with a poorly characterized physiological role in regulating the synaptic vesicle cycle. Using RBL-2H3 cells as a model system, we earlier reported that wild-type alpha-synuclein can act as both an inhibitor and a potentiator of stimulated exocytosis in a concentration-dependent manner. The inhibitory function is constitutive and depends on membrane binding by the helix-2 region of the lipid-binding domain, while potentiation becomes apparent only at high concentrations. Using structural and functional characterization of conformationally selective mutants via a combination of spectroscopic and cellular assays, we show here that binding affinity for isolated vesicles similar in size to synaptic vesicles is a primary determinant of alpha-synuclein-mediated potentiation of vesicle release. Inhibition of release is sensitive to changes in the region linking the helix-1 and helix-2 regions of the N-terminal lipid-binding domain and may require some degree of coupling between these regions. Potentiation of release likely occurs as a result of alpha-synuclein interactions with undocked vesicles isolated away from the active zone in internal pools. Consistent with this, we observe that alpha-synuclein can disperse vesicles from in vitro clusters organized by condensates of the presynaptic protein synapsin-1.
Keywords: Parkinson’s; alpha-synuclein; membrane; synapsin; synaptic vesicle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The docking of synaptic vesicles on the presynaptic membrane induced by α-synuclein is modulated by lipid composition.Nat Commun. 2021 Feb 10;12(1):927. doi: 10.1038/s41467-021-21027-4. Nat Commun. 2021. PMID: 33568632 Free PMC article.
-
The physiological role of α-synuclein and its relationship to Parkinson's Disease.J Neurochem. 2019 Sep;150(5):475-486. doi: 10.1111/jnc.14810. Epub 2019 Jul 28. J Neurochem. 2019. PMID: 31269263 Free PMC article. Review.
-
Acute increase of α-synuclein inhibits synaptic vesicle recycling evoked during intense stimulation.Mol Biol Cell. 2014 Dec 1;25(24):3926-41. doi: 10.1091/mbc.E14-02-0708. Epub 2014 Oct 1. Mol Biol Cell. 2014. PMID: 25273557 Free PMC article.
-
Synaptic vesicle binding of α-synuclein is modulated by β- and γ-synucleins.Cell Rep. 2022 Apr 12;39(2):110675. doi: 10.1016/j.celrep.2022.110675. Cell Rep. 2022. PMID: 35417693 Free PMC article.
-
Functional and Pathological Effects of α-Synuclein on Synaptic SNARE Complexes.J Mol Biol. 2023 Jan 15;435(1):167714. doi: 10.1016/j.jmb.2022.167714. Epub 2022 Jul 3. J Mol Biol. 2023. PMID: 35787839 Free PMC article. Review.
Cited by
-
Alpha synuclein modulates mitochondrial Ca2+ uptake from ER during cell stimulation and under stress conditions.NPJ Parkinsons Dis. 2023 Sep 23;9(1):137. doi: 10.1038/s41531-023-00578-x. NPJ Parkinsons Dis. 2023. PMID: 37741841 Free PMC article.
-
Per Aspera ad Chaos: Vladimir Uversky's Odyssey through the Strange World of Intrinsically Disordered Proteins.Biomolecules. 2023 Jun 19;13(6):1015. doi: 10.3390/biom13061015. Biomolecules. 2023. PMID: 37371595 Free PMC article.
-
Exploring the Application Potential of α-Synuclein Molecular Probes in Early Diagnosis of Parkinson's Disease: Focus on Imaging Methods.ACS Chem Neurosci. 2025 May 21;16(10):1838-1846. doi: 10.1021/acschemneuro.5c00008. Epub 2025 May 7. ACS Chem Neurosci. 2025. PMID: 40334069 Free PMC article. Review.
-
Recent advances in nanotechnology for Parkinson's disease: diagnosis, treatment, and future perspectives.Front Med (Lausanne). 2025 Jan 22;12:1535682. doi: 10.3389/fmed.2025.1535682. eCollection 2025. Front Med (Lausanne). 2025. PMID: 39911864 Free PMC article. Review.
-
Non-uniform sampling of similar NMR spectra and its application to studies of the interaction between alpha-synuclein and liposomes.J Biomol NMR. 2023 Aug;77(4):149-163. doi: 10.1007/s10858-023-00418-3. Epub 2023 May 26. J Biomol NMR. 2023. PMID: 37237169 Free PMC article.
References
-
- Feigin V.L., Nichols E., Alam T., Bannick M.S., Beghi E., Blake N., Culpepper W.J., Dorsey E.R., Elbaz A., Ellenbogen R.G., et al. Global, Regional, and National Burden of Neurological Disorders, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019;18:459–480. doi: 10.1016/S1474-4422(18)30499-X. - DOI - PMC - PubMed
-
- Mahul-Mellier A.-L., Burtscher J., Maharjan N., Weerens L., Croisier M., Kuttler F., Leleu M., Knott G.W., Lashuel H.A. The Process of Lewy Body Formation, Rather than Simply α-Synuclein Fibrillization, Is One of the Major Drivers of Neurodegeneration. Proc. Natl. Acad. Sci. USA. 2020;117:4971–4982. doi: 10.1073/pnas.1913904117. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R35 GM136686/GM/NIGMS NIH HHS/United States
- R01 GM117552/GM/NIGMS NIH HHS/United States
- S10 OD016432/OD/NIH HHS/United States
- MCB-1950525/National Science Foundation
- C06 RR015495/RR/NCRR NIH HHS/United States
- RM1 GM145397/GM/NIGMS NIH HHS/United States
- S10 RR025502/RR/NCRR NIH HHS/United States
- S10 OD016320/OD/NIH HHS/United States
- P41 GM066354/GM/NIGMS NIH HHS/United States
- P41 GM118302/GM/NIGMS NIH HHS/United States
- S10 OD018509/OD/NIH HHS/United States
- R37 AG019391/AG/NIA NIH HHS/United States
- 1428922/National Science Foundation
LinkOut - more resources
Full Text Sources
Medical
