

iPSC-Derived Macrophages: The Differentiation Protocol Affects Cell Immune Characteristics and Differentiation Trajectories
- PMID: 36555728
- PMCID: PMC9781144
- DOI: 10.3390/ijms232416087
iPSC-Derived Macrophages: The Differentiation Protocol Affects Cell Immune Characteristics and Differentiation Trajectories
Abstract
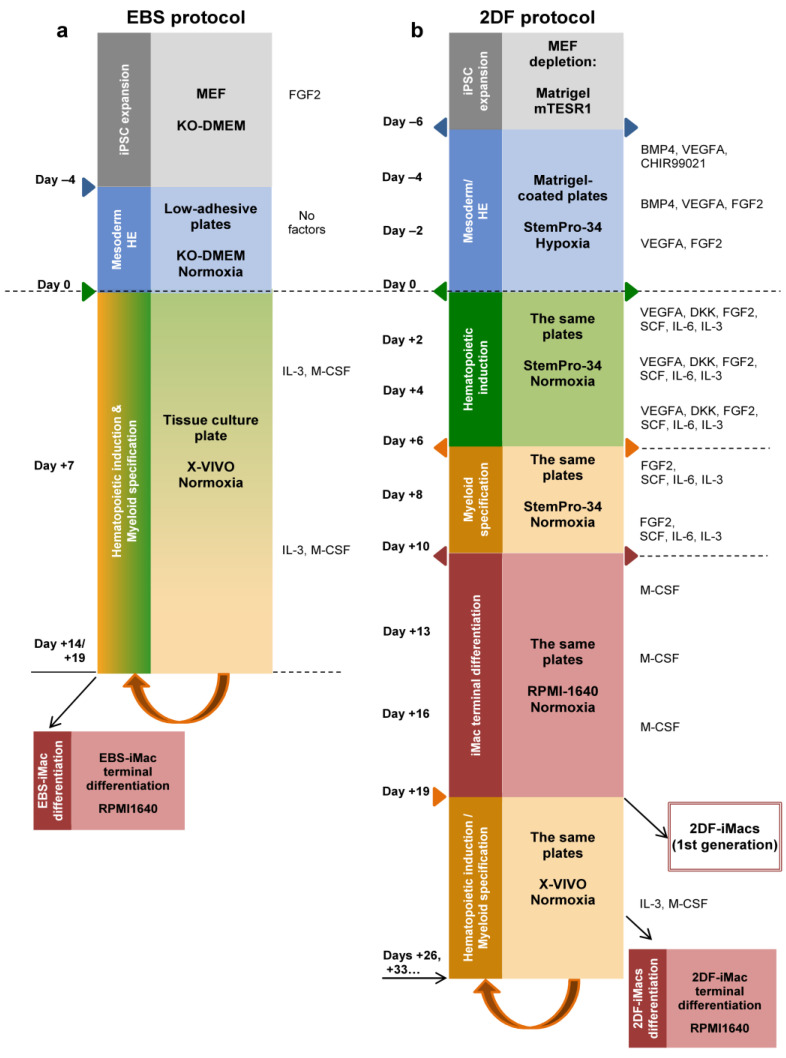
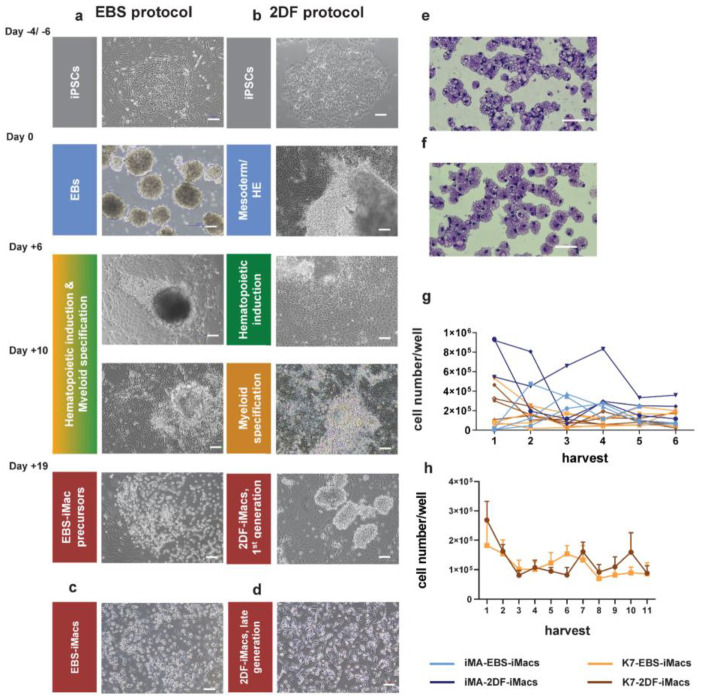
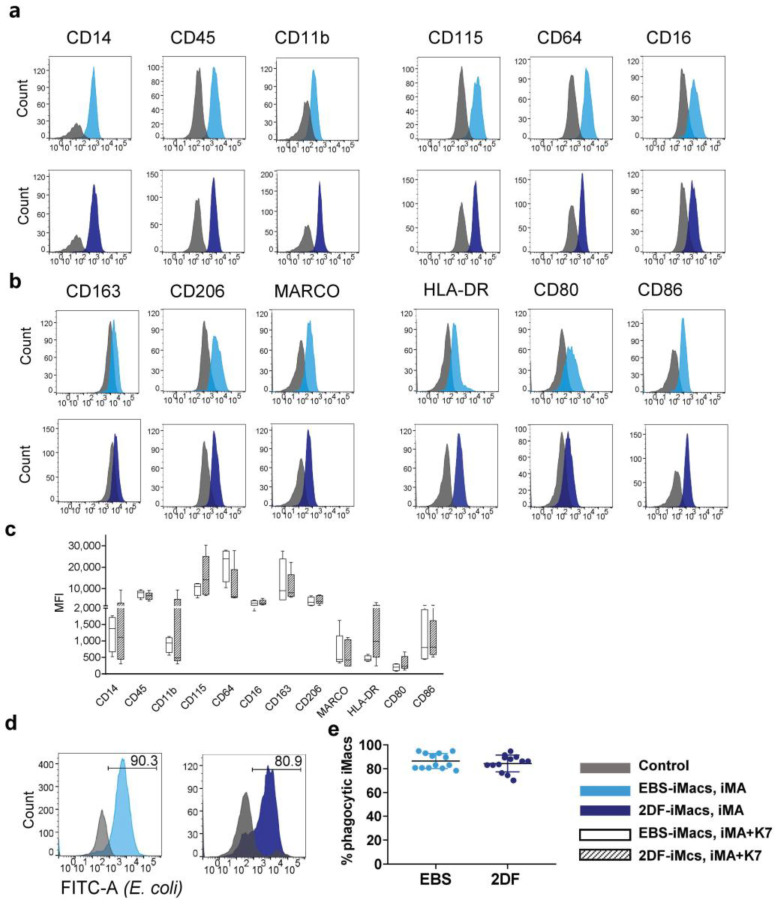
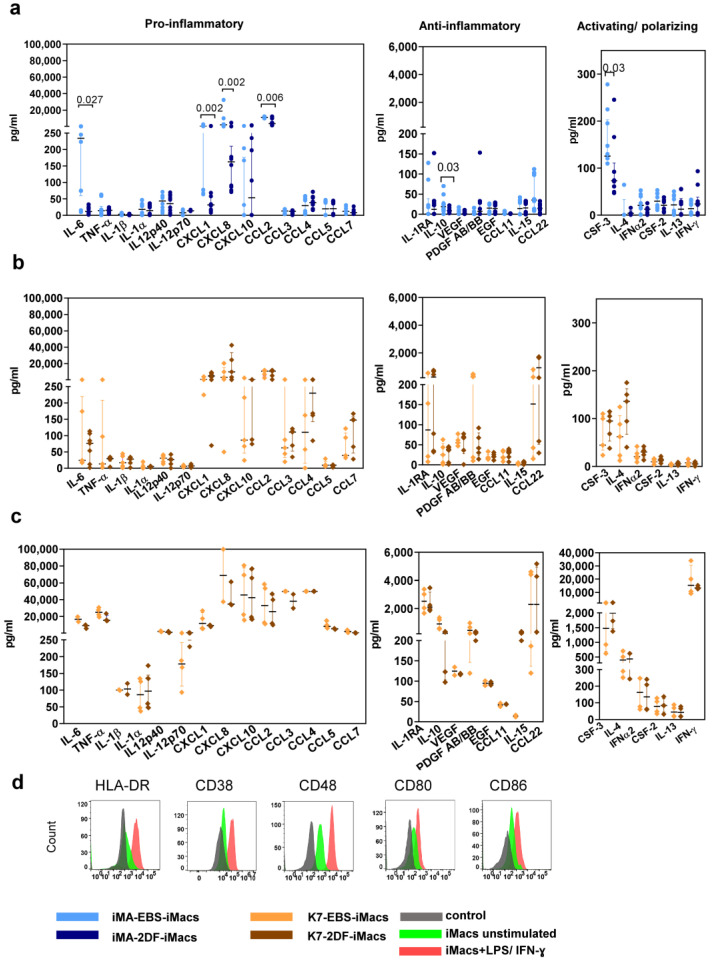
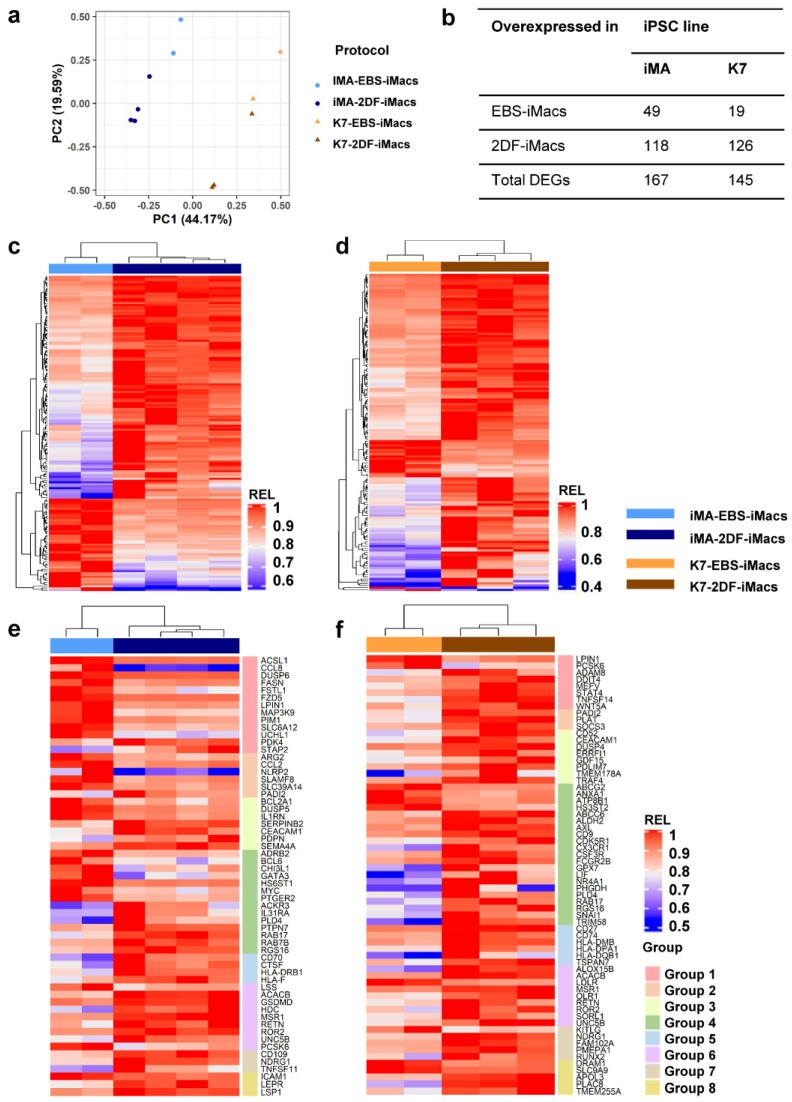
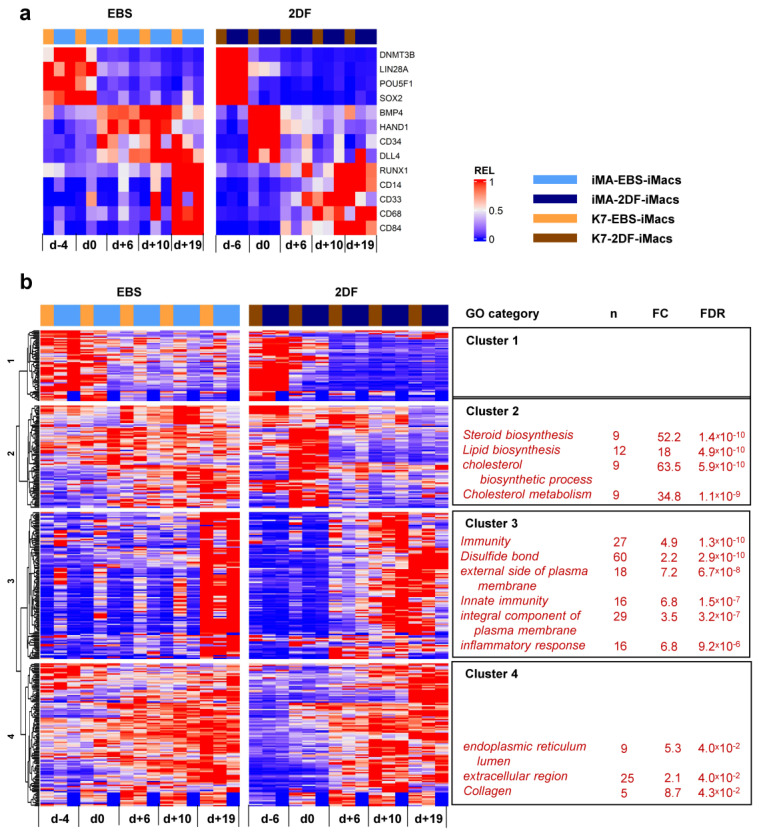
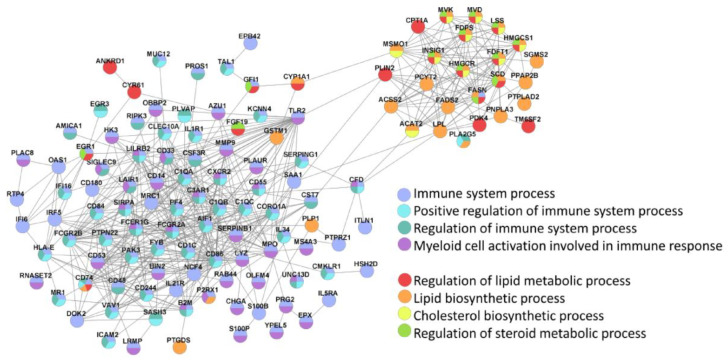
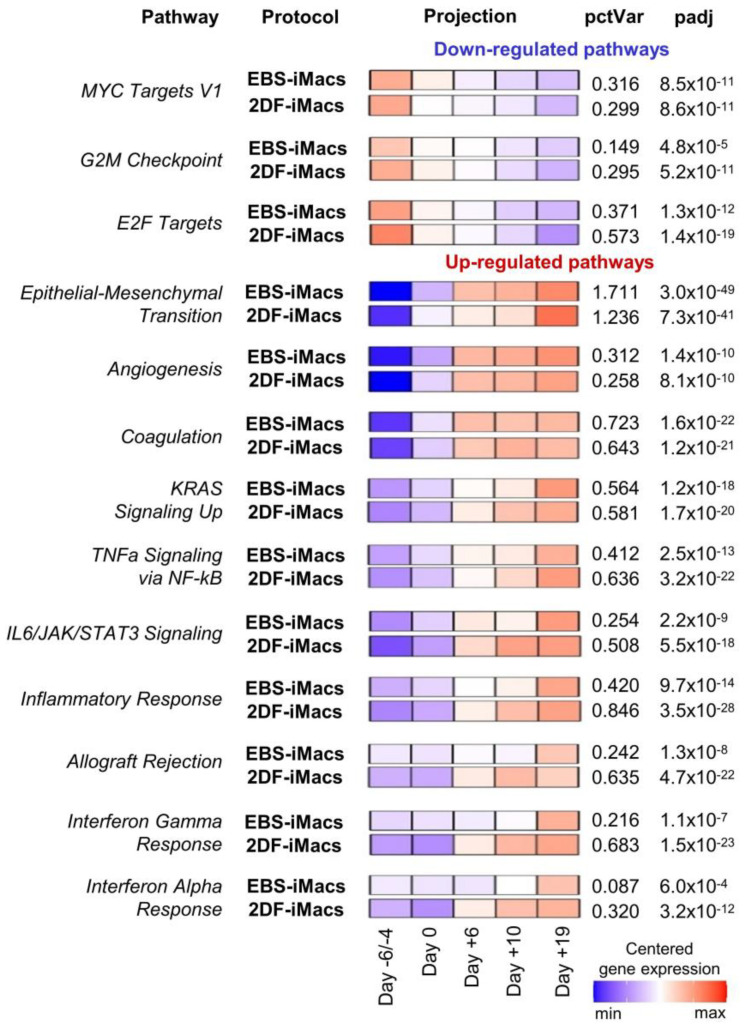
The generation of human macrophages from induced pluripotent stem cells (iMacs) is a rapidly developing approach used to create disease models, screen drugs, study macrophage-pathogen interactions and develop macrophage-based cell therapy. To generate iMacs, different types of protocols have been suggested, all thought to result in the generation of similar iMac populations. However, direct comparison of iMacs generated using different protocols has not been performed. We have compared the productivity, the differentiation trajectories and the characteristics of iMacs generated using two widely used protocols: one based on the formation of embryoid bodies and the induction of myeloid differentiation by only two cytokines, interleukin-3 and macrophage colony-stimulating factor, and the other utilizing multiple exogenous factors for iMac generation. We report inter-protocol differences in the following: (i) protocol productivity; (ii) dynamic changes in the expression of genes related to inflammation and lipid homeostasis following iMac differentiation and (iii) the transcriptomic profiles of terminally differentiated iMacs, including the expression of genes involved in inflammatory response, antigen presentation and lipid homeostasis. The results document the dependence of fine iMac characteristics on the type of differentiation protocol, which is important for further development of the field, including the development of iMac-based cell therapy.
Keywords: antigen presentation; induced pluripotent stem cells; inflammatory response; lipid homeostasis; macrophage differentiation; macrophages derived from induced pluripotent stem cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Panicker L.M., Miller D., Park T.S., Pate B., Azevedo J.L., Awad O., Masood M.A., Veenstra T.D., Goldin E., Stubblefield B.K., et al. Induced pluripotent stem cell model recapitulates pathologic hallmarks of Gaucher disease. Proc. Natl. Acad. Sci. USA. 2012;109:18054–18059. doi: 10.1073/pnas.1207889109. - DOI - PMC - PubMed
-
- Brault J., Goutagny E., Telugu N., Shao K., Baquié M., Satre V., Coutton C., Grunwald D., Brion J.P., Barlogis V., et al. Optimized generation of functional neutrophils and macrophages from patient-specific induced pluripotent stem cells: Ex vivo models of X(0)-linked, AR22(0)- and AR47(0)- chronic granulomatous diseases. BioResearch. 2014;3:311–326. doi: 10.1089/biores.2014.0045. - DOI - PMC - PubMed
-
- Zhang H., Xue C., Shah R., Bermingham K., Hinkle C.C., Li W., Rodrigues A., Tabita-Martinez J., Millar J.S., Cuchel M., et al. Functional analysis and transcriptomic profiling of iPSC-derived macrophages and their application in modeling Mendelian disease. Circ. Res. 2015;117:17–28. doi: 10.1161/CIRCRESAHA.117.305860. - DOI - PMC - PubMed
-
- Klatt D., Cheng E., Philipp F., Selich A., Dahlke J., Schmidt R.E., Schott J.W., Büning H., Hoffmann D., Thrasher A.J., et al. Targeted repair of p47-CGD in iPSCs by CRISPR/Cas9: Functional correction without cleavage in the highly homologous pseudogenes. Stem Cell Rep. 2019;13:590–598. doi: 10.1016/j.stemcr.2019.08.008. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases