

Cell-Type-Dependent Role for nsP3 Macrodomain ADP-Ribose Binding and Hydrolase Activity during Chikungunya Virus Infection
- PMID: 36560748
- PMCID: PMC9787352
- DOI: 10.3390/v14122744
Cell-Type-Dependent Role for nsP3 Macrodomain ADP-Ribose Binding and Hydrolase Activity during Chikungunya Virus Infection
Abstract
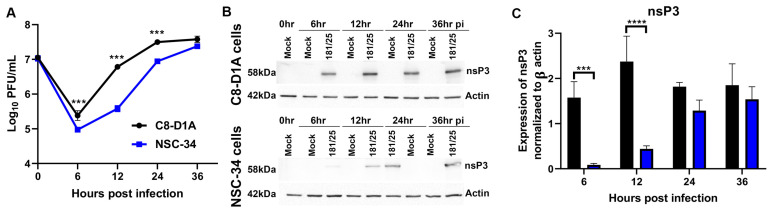
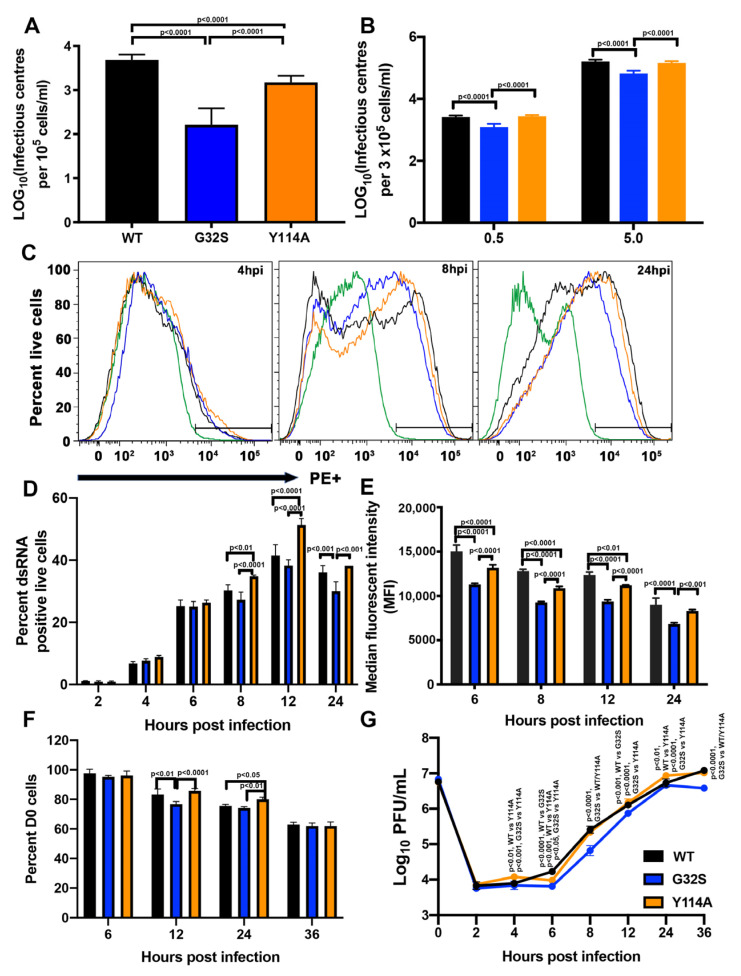
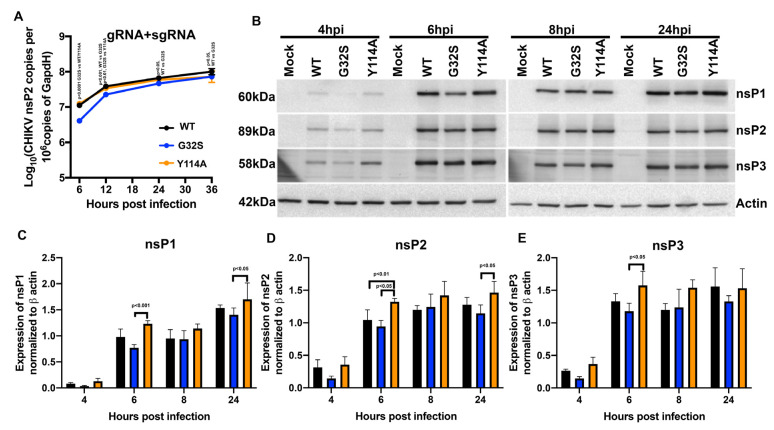
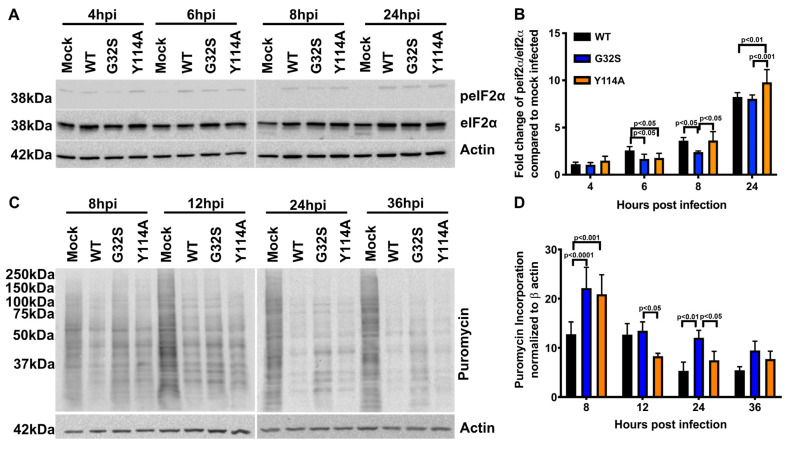
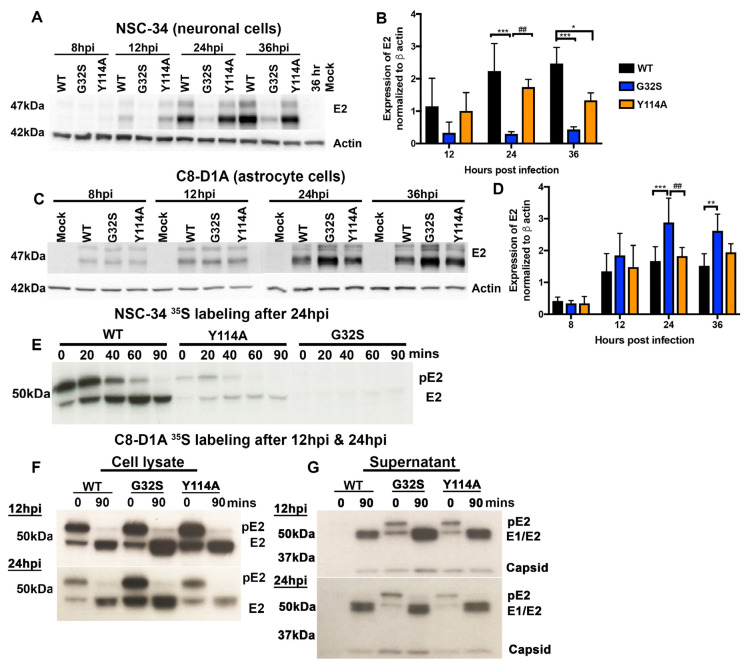
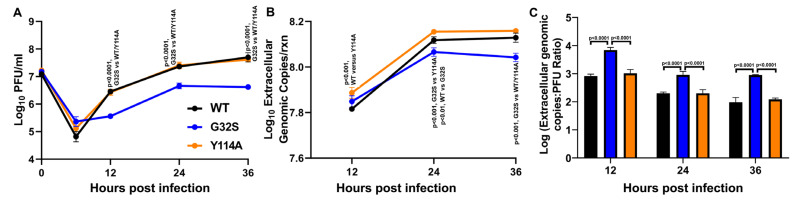
Chikungunya virus (CHIKV) causes outbreaks of rash, arthritis, and fever associated with neurologic complications, where astrocytes are preferentially infected. A determinant of virulence is the macrodomain (MD) of nonstructural protein 3 (nsP3), which binds and removes ADP-ribose (ADPr) from ADP-ribosylated substrates and regulates stress-granule disruption. We compared the replication of CHIKV 181/25 (WT) and MD mutants with decreased ADPr binding and hydrolase (G32S) or increased ADPr binding and decreased hydrolase (Y114A) activities in C8-D1A astrocytic cells and NSC-34 neuronal cells. WT CHIKV replication was initiated more rapidly with earlier nsP synthesis in C8-D1A than in NSC-34 cells. G32S established infection, amplified replication complexes, and induced host-protein synthesis shut-off less efficiently than WT and produced less infectious virus, while Y114A replication was close to WT. However, G32S mutation effects on structural protein synthesis were cell-type-dependent. In NSC-34 cells, E2 synthesis was decreased compared to WT, while in C8-D1A cells synthesis was increased. Excess E2 produced by G32S-infected C8-D1A cells was assembled into virus particles that were less infectious than those from WT or Y114A-infected cells. Because nsP3 recruits ADP-ribosylated RNA-binding proteins in stress granules away from translation-initiation factors into nsP3 granules where the MD hydrolase can remove ADPr, we postulate that suboptimal translation-factor release decreased structural protein synthesis in NSC-34 cells while failure to de-ADP-ribosylate regulatory RNA-binding proteins increased synthesis in C8-D1A cells.
Keywords: ADP ribosylation; alphavirus; astrocytes; chikungunya virus; host-virus interaction; macrodomain; neurons; neurovirulence; replication complexes; viral replication.
Conflict of interest statement
J.X.Y. is an employee of Takeda Pharmaceuticals. D.E.G. is a member of the GlaxoSmithKline Vaccines Research and Development Advisory Board, the Takeda Pharmaceuticals Zika virus Vaccine Data Monitoring Committee, Academia Sinica Genome Research Center Advisory Board, and GreenLight Biosciences Vaccine Development Scientific Advisory Board. T.K., R.A. and L.P. declare no competing interest.
Figures
References
-
- Hucke F.I.L., Bestehorn-Willmann M., Bassetto M., Brancale A., Zanetta P., Bugert J.J. CHIKV strains Brazil (wt) and Ross (lab-adapted) differ with regard to cell host range and antiviral sensitivity and show CPE in human glioblastoma cell lines U138 and U251. Virus Genes. 2022;58:188–202. doi: 10.1007/s11262-022-01892-x. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
