

Widespread, Reversible Cysteine Modification by Methylglyoxal Regulates Metabolic Enzyme Function
- PMID: 36562291
- PMCID: PMC9872086
- DOI: 10.1021/acschembio.2c00727
Widespread, Reversible Cysteine Modification by Methylglyoxal Regulates Metabolic Enzyme Function
Abstract
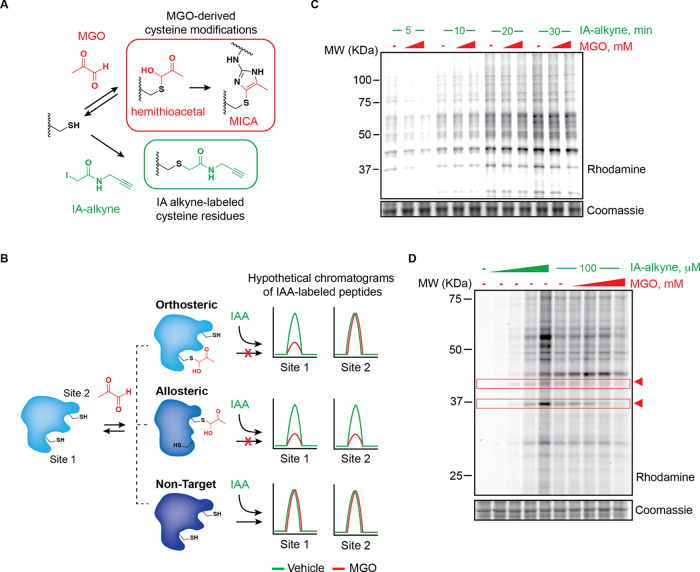
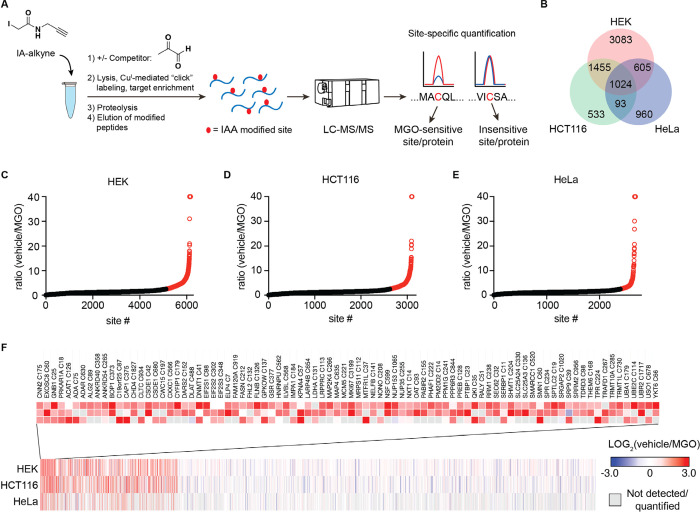
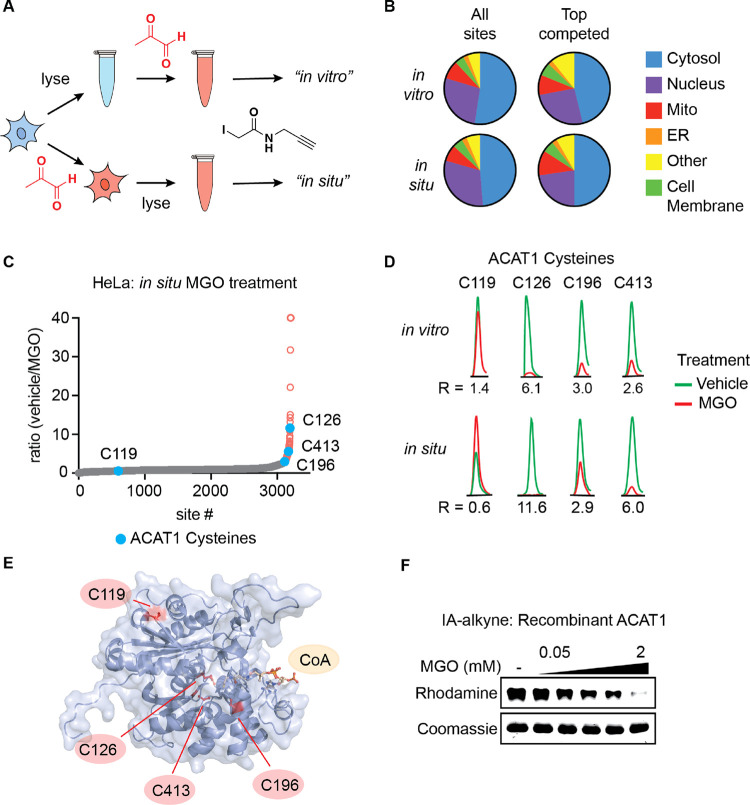
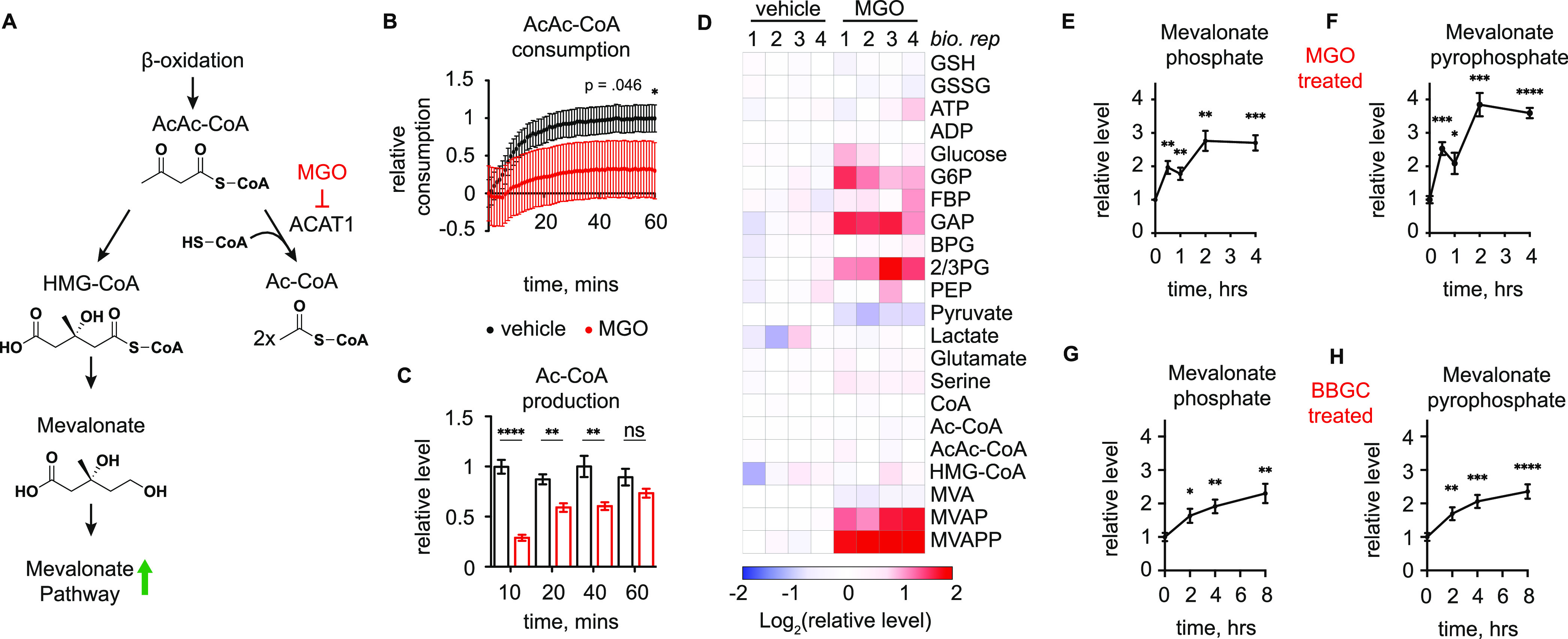
Methylglyoxal (MGO), a reactive metabolite byproduct of glucose metabolism, is known to form a variety of posttranslational modifications (PTMs) on nucleophilic amino acids. For example, cysteine, the most nucleophilic proteinogenic amino acid, forms reversible hemithioacetal and stable mercaptomethylimidazole adducts with MGO. The high reactivity of cysteine toward MGO and the rate of formation of such modifications provide the opportunity for mechanisms by which proteins and pathways might rapidly sense and respond to alterations in levels of MGO. This indirect measure of alterations in glycolytic flux would thereby allow disparate cellular processes to dynamically respond to changes in nutrient availability and utilization. Here we report the use of quantitative LC-MS/MS-based chemoproteomic profiling approaches with a cysteine-reactive probe to map the proteome-wide landscape of MGO modification of cysteine residues. This approach led to the identification of many sites of potential functional regulation by MGO. We further characterized the role that such modifications have in a catalytic cysteine residue in a key metabolic enzyme and the resulting effects on cellular metabolism.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Salomón T.; Sibbersen C.; Hansen J.; Britz D.; Svart M. V.; Voss T. S.; Møller N.; Gregersen N.; Jørgensen K. A.; Palmfeldt J.; Poulsen T. B.; Johannsen M. Ketone Body Acetoacetate Buffers Methylglyoxal via a Non-enzymatic Conversion during Diabetic and Dietary Ketosis. Cell Chem. Biol. 2017, 24, 935–943. 10.1016/j.chembiol.2017.07.012. - DOI - PubMed
-
- Henle T. W.; Walter A. W.; Haeßner R.; Klostermeyer H. Detection and identification of a protein-bound imidazolone resulting from the reaction of arginine residues and methylglyoxal. Z. Lebensm.-Unters Forsch. 1994, 199, 55–58. 10.1007/BF01192954. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
