

Mobile genetic elements from the maternal microbiome shape infant gut microbial assembly and metabolism
- PMID: 36563663
- PMCID: PMC9869402
- DOI: 10.1016/j.cell.2022.11.023
Mobile genetic elements from the maternal microbiome shape infant gut microbial assembly and metabolism
Abstract
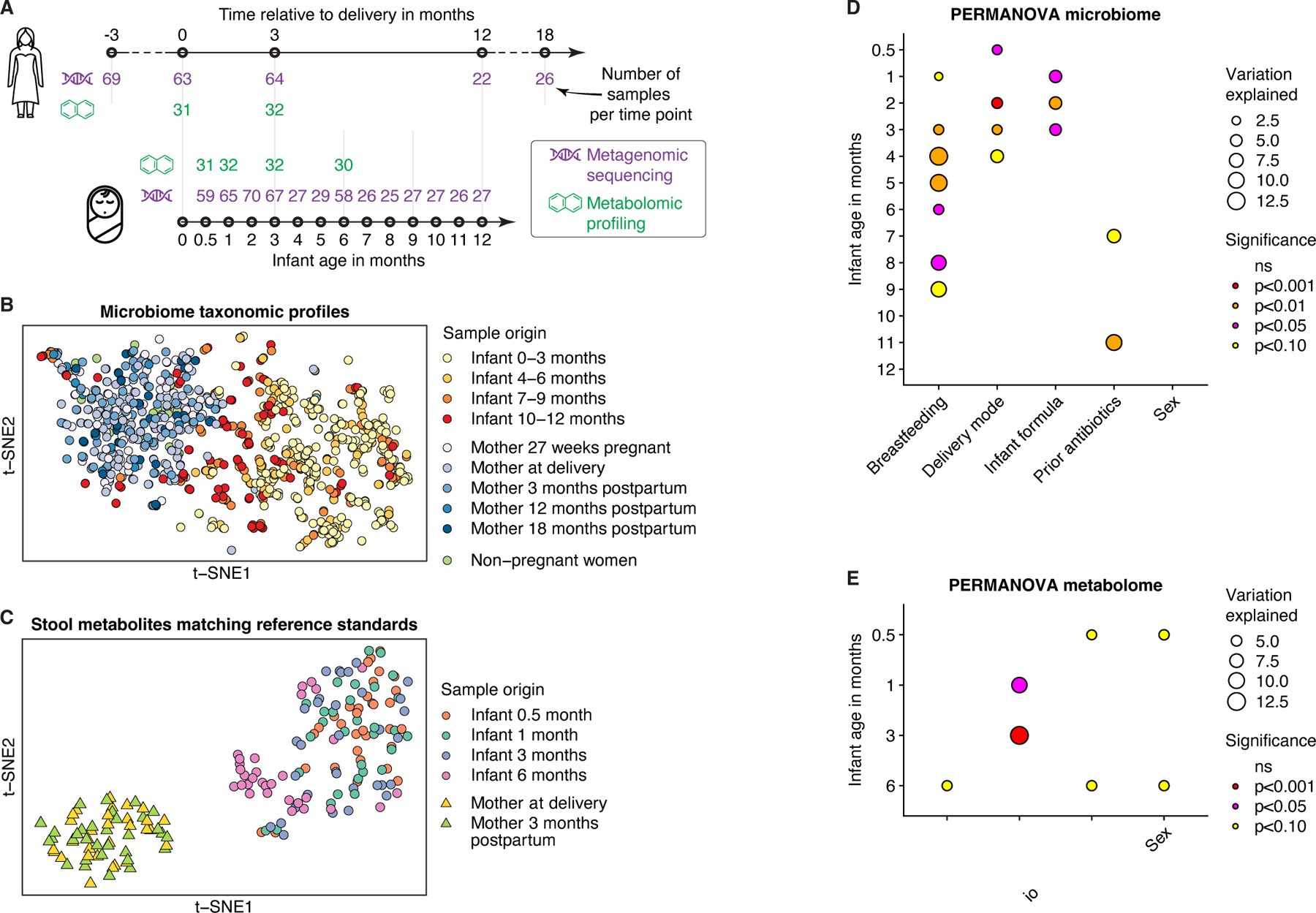
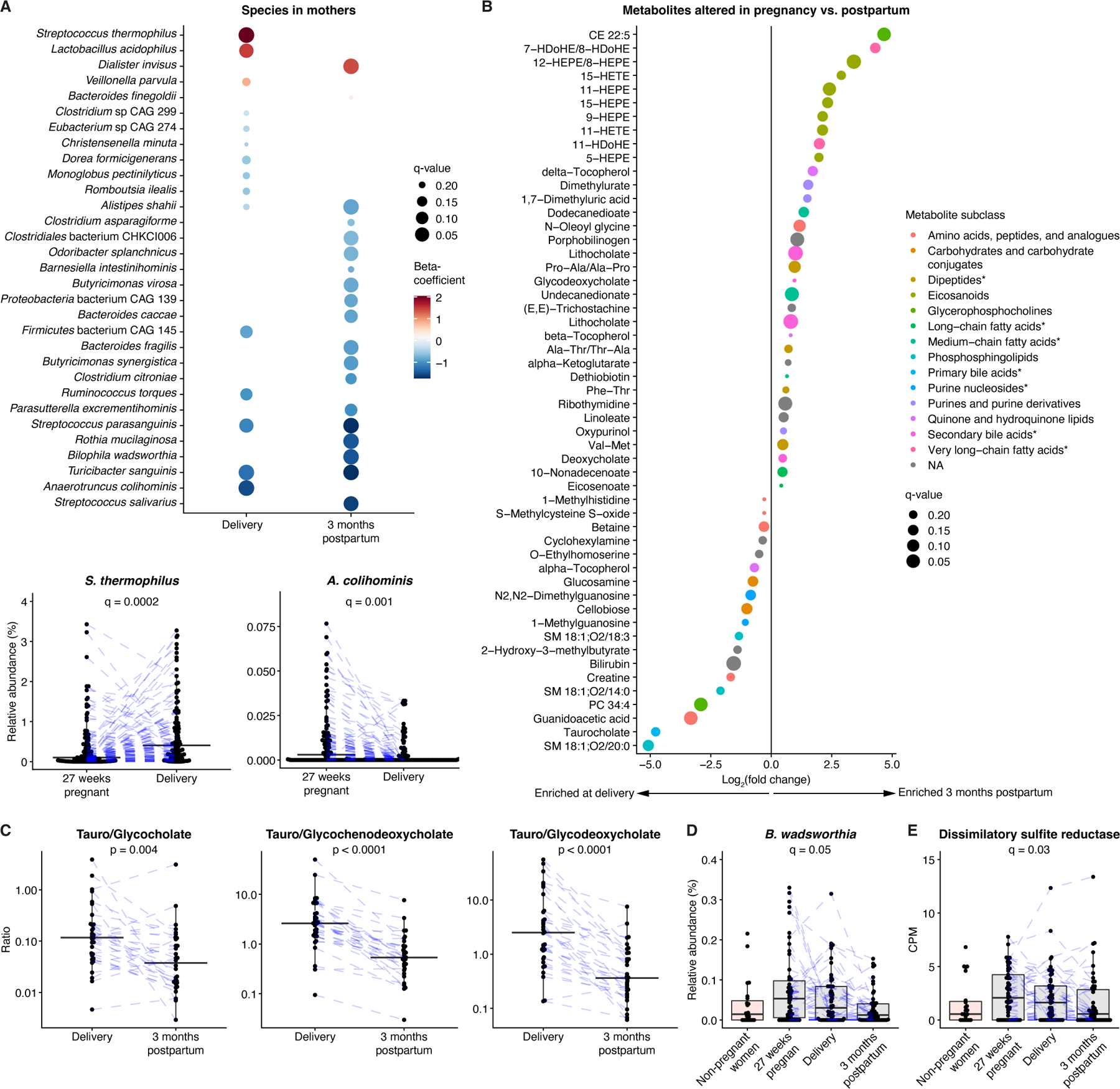
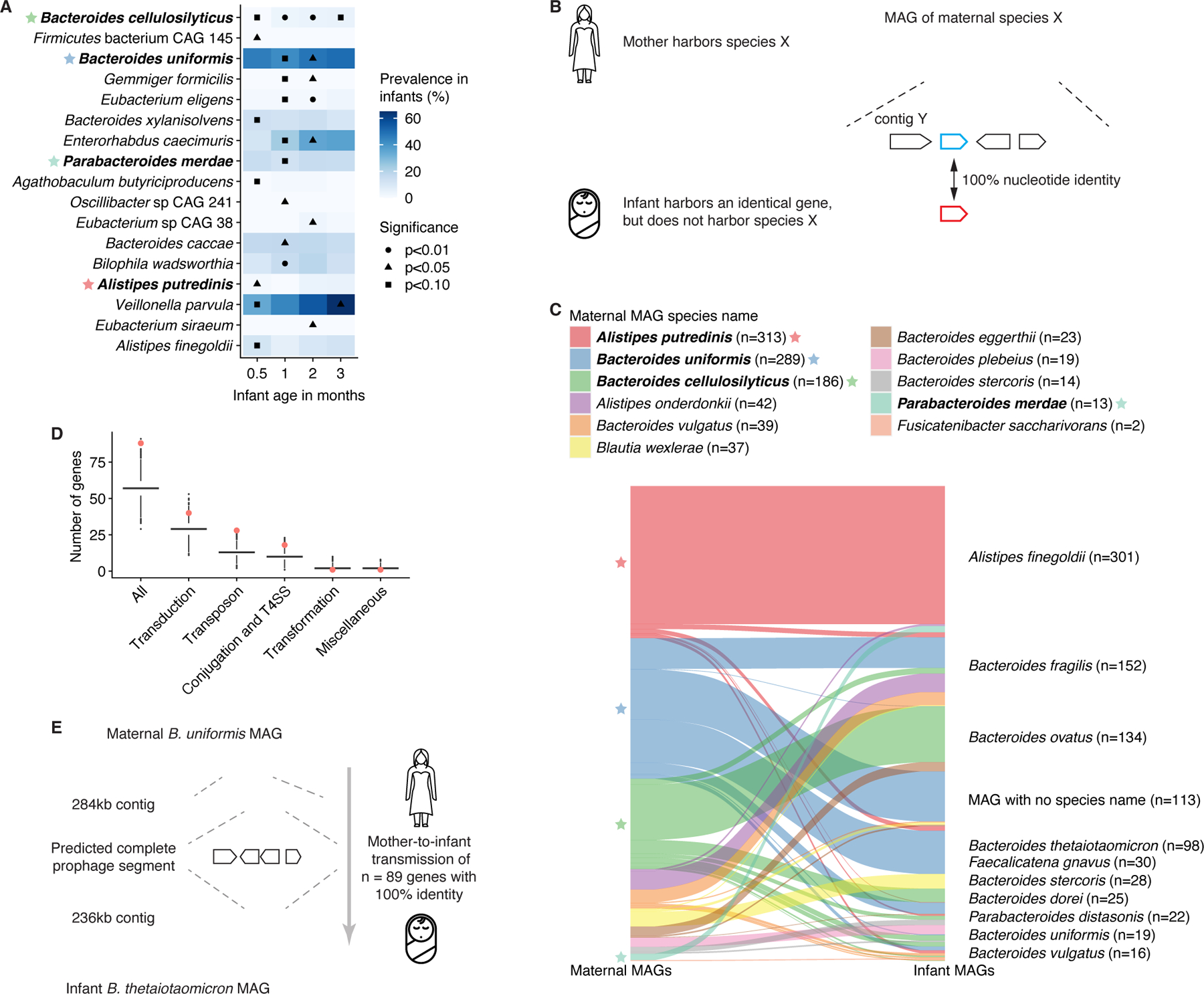
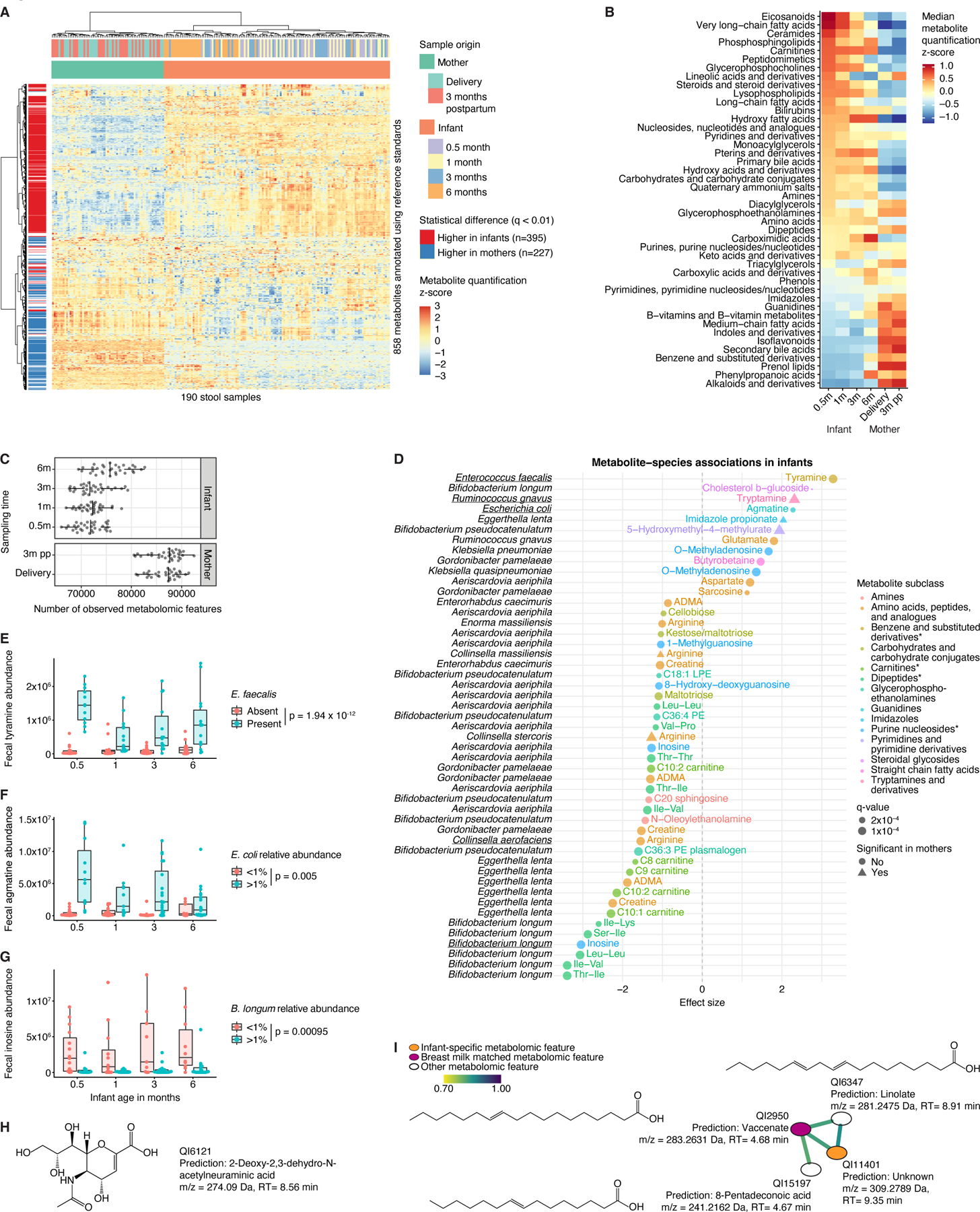
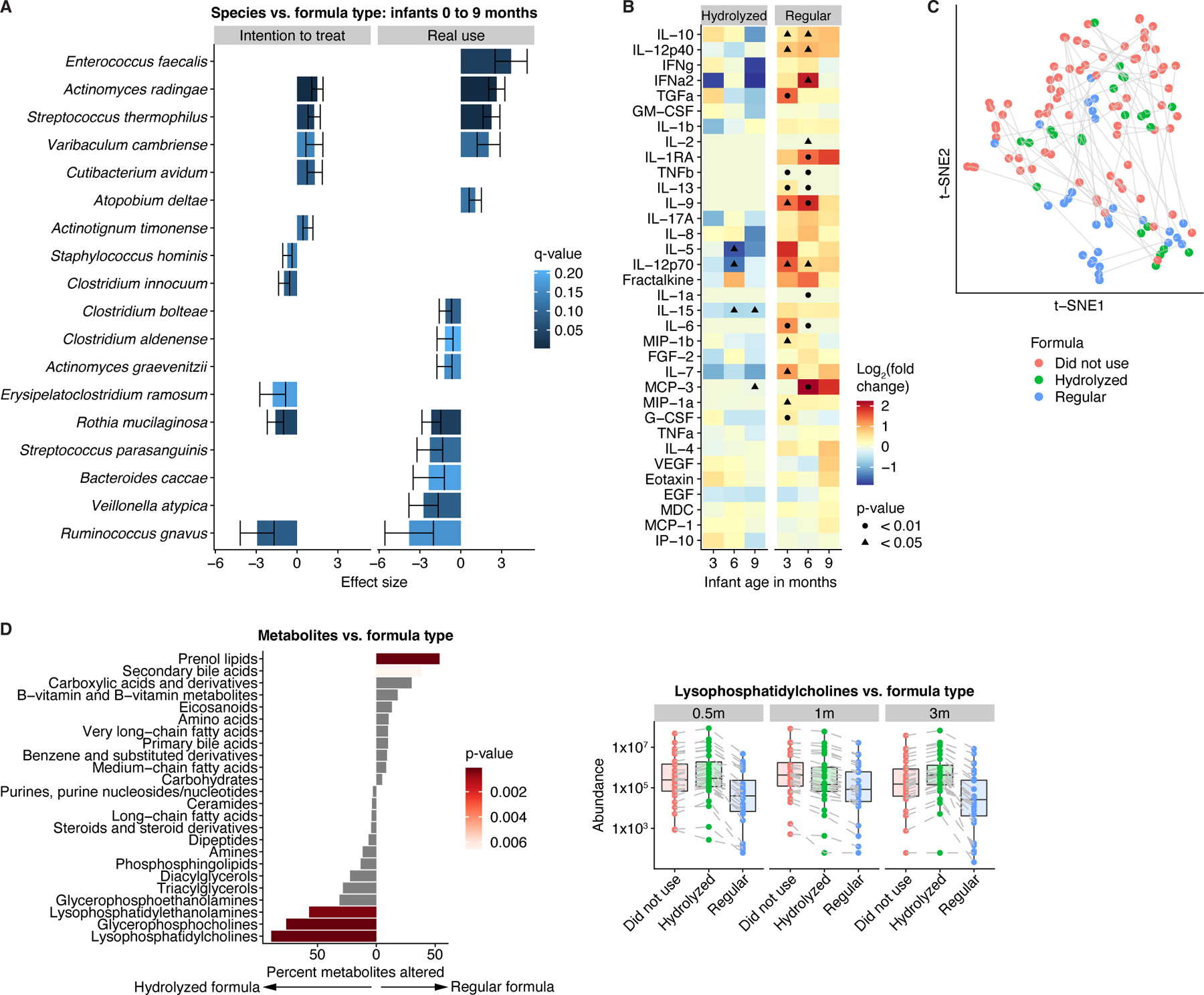
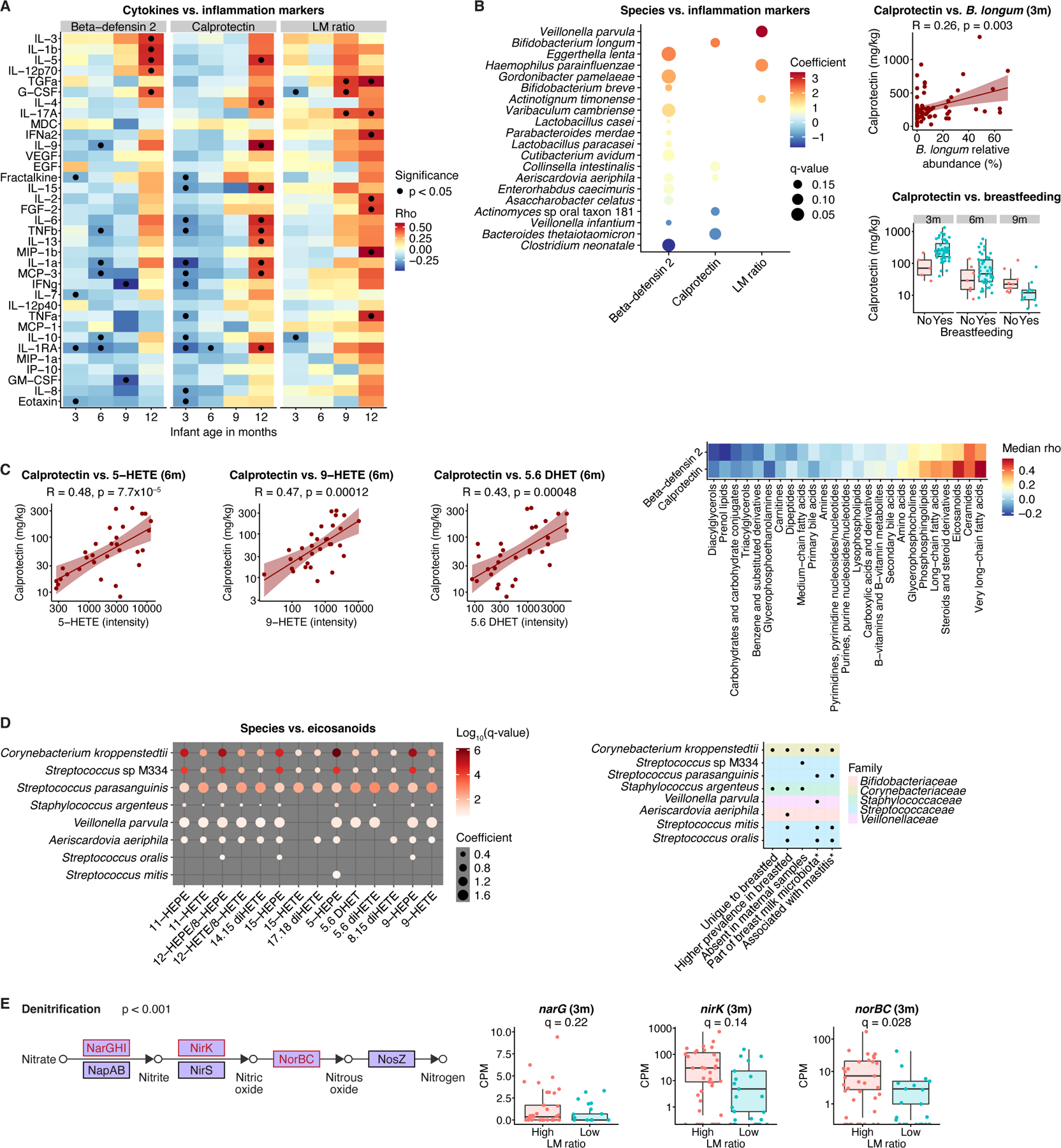
The perinatal period represents a critical window for cognitive and immune system development, promoted by maternal and infant gut microbiomes and their metabolites. Here, we tracked the co-development of microbiomes and metabolomes from late pregnancy to 1 year of age using longitudinal multi-omics data from a cohort of 70 mother-infant dyads. We discovered large-scale mother-to-infant interspecies transfer of mobile genetic elements, frequently involving genes associated with diet-related adaptations. Infant gut metabolomes were less diverse than maternal but featured hundreds of unique metabolites and microbe-metabolite associations not detected in mothers. Metabolomes and serum cytokine signatures of infants who received regular-but not extensively hydrolyzed-formula were distinct from those of exclusively breastfed infants. Taken together, our integrative analysis expands the concept of vertical transmission of the gut microbiome and provides original insights into the development of maternal and infant microbiomes and metabolomes during late pregnancy and early life.
Keywords: gut metabolome; horizontal gene transfer; infant gut microbiome; mother-to-infant microbiome transmission.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests R.J.X. is co-founder of Jnana Therapeutics and Celsius Therapeutics, board director at MoonLake Immunotherapeutics, and consultant to Nestlé; these organizations had no role in the study.
Figures
References
-
- Derrien M, Alvarez A-S, and de Vos WM (2019). The Gut Microbiota in the First Decade of Life. Trends Microbiol 27, 997–1010. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
