

Substrate Specificity and Kinetics of RNA Hydrolysis by SARS-CoV-2 NSP10/14 Exonuclease
- PMID: 36570070
- PMCID: PMC9718090
- DOI: 10.1021/acsbiomedchemau.2c00046
Substrate Specificity and Kinetics of RNA Hydrolysis by SARS-CoV-2 NSP10/14 Exonuclease
Abstract
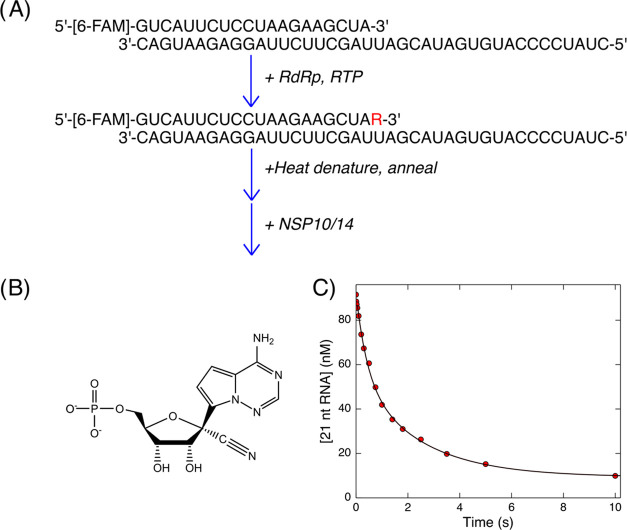
Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), the virus that causes COVID-19, continues to evolve resistance to vaccines and existing antiviral therapies at an alarming rate, increasing the need for new direct-acting antiviral drugs. Despite significant advances in our fundamental understanding of the kinetics and mechanism of viral RNA replication, there are still open questions regarding how the proofreading exonuclease (NSP10/NSP14 complex) contributes to replication fidelity and resistance to nucleoside analogs. Through single turnover kinetic analysis, we show that the preferred substrate for the exonuclease is double-stranded RNA without any mismatches. Double-stranded RNA containing a 3'-terminal remdesivir was hydrolyzed at a rate similar to a correctly base-paired cognate nucleotide. Surprisingly, single-stranded RNA or duplex RNA containing a 3'-terminal mismatch was hydrolyzed at rates 125- and 45-fold slower, respectively, compared to the correctly base-paired double-stranded RNA. These results define the substrate specificity and rate of removal of remdesivir for the exonuclease and outline rigorous kinetic assays that could help in finding next-generation exonuclease inhibitors or nucleoside analogs that are able to evade excision. These results also raise important questions about the role of the polymerase/exonuclease complex in proofreading during viral replication. Addressing these questions through rigorous kinetic analysis will facilitate the search for desperately needed antiviral drugs to combat COVID-19.
© 2022 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare the following competing financial interest(s): K.A.J. is president of KinTek Corporation, which provided the RQF-3 rapid quench flow instrument and KinTek Explorer software used in this study.
Figures


Similar articles
-
Activity of nsp14 Exonuclease from SARS-CoV-2 towards RNAs with Modified 3'-Termini.Dokl Biochem Biophys. 2023 Apr;509(1):65-69. doi: 10.1134/S1607672923700102. Epub 2023 Jun 20. Dokl Biochem Biophys. 2023. PMID: 37340295
-
Reconstitution and functional characterization of SARS-CoV-2 proofreading complex.Protein Expr Purif. 2021 Sep;185:105894. doi: 10.1016/j.pep.2021.105894. Epub 2021 Apr 29. Protein Expr Purif. 2021. PMID: 33933612 Free PMC article.
-
Mechanisms of inhibition of viral RNA replication by nucleotide analogs.Enzymes. 2021;49:39-62. doi: 10.1016/bs.enz.2021.07.001. Epub 2021 Sep 27. Enzymes. 2021. PMID: 34696838 Free PMC article. Review.
-
The coronavirus nsp14 exoribonuclease interface with the cofactor nsp10 is essential for efficient virus replication and enzymatic activity.bioRxiv [Preprint]. 2024 Sep 26:2024.09.26.615217. doi: 10.1101/2024.09.26.615217. bioRxiv. 2024. Update in: J Virol. 2025 Feb 25;99(2):e0170824. doi: 10.1128/jvi.01708-24. PMID: 39386528 Free PMC article. Updated. Preprint.
-
Opportunities and Challenges in Targeting the Proofreading Activity of SARS-CoV-2 Polymerase Complex.Molecules. 2022 May 3;27(9):2918. doi: 10.3390/molecules27092918. Molecules. 2022. PMID: 35566268 Free PMC article. Review.
Cited by
-
Adenosine modifications impede SARS-CoV-2 RNA-dependent RNA transcription.RNA. 2024 Aug 16;30(9):1141-1150. doi: 10.1261/rna.079991.124. RNA. 2024. PMID: 38942480 Free PMC article.
References
-
- Baddock H. T.; Brolih S.; Yosaatmadja Y.; Ratnaweera M.; Bielinski M.; Swift; Lonnie P.; Cruz-Migoni A.; Fan H.; Keown J. R.; Walker A. P.; et al. Characterization of the SARS-CoV-2 ExoN (nsp14ExoN–nsp10) complex: implications for its role in viral genome stability and inhibitor identification. Nucleic Acids Res. 2022, 50, 1484–1500. 10.1093/nar/gkab1303. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
