

Diverse monogenic subforms of human spermatogenic failure
- PMID: 36572685
- PMCID: PMC9792524
- DOI: 10.1038/s41467-022-35661-z
Diverse monogenic subforms of human spermatogenic failure
Abstract
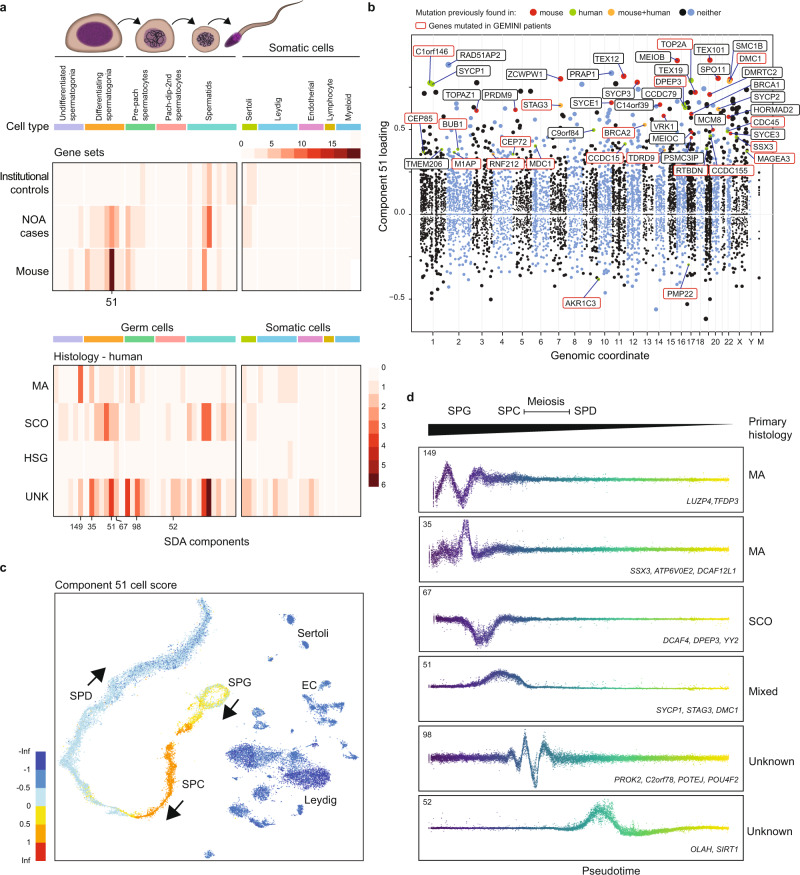
Non-obstructive azoospermia (NOA) is the most severe form of male infertility and typically incurable. Defining the genetic basis of NOA has proven challenging, and the most advanced classification of NOA subforms is not based on genetics, but simple description of testis histology. In this study, we exome-sequenced over 1000 clinically diagnosed NOA cases and identified a plausible recessive Mendelian cause in 20%. We find further support for 21 genes in a 2-stage burden test with 2072 cases and 11,587 fertile controls. The disrupted genes are primarily on the autosomes, enriched for undescribed human "knockouts", and, for the most part, have yet to be linked to a Mendelian trait. Integration with single-cell RNA sequencing data shows that azoospermia genes can be grouped into molecular subforms with synchronized expression patterns, and analogs of these subforms exist in mice. This analysis framework identifies groups of genes with known roles in spermatogenesis but also reveals unrecognized subforms, such as a set of genes expressed across mitotic divisions of differentiating spermatogonia. Our findings highlight NOA as an understudied Mendelian disorder and provide a conceptual structure for organizing the complex genetics of male infertility, which may provide a rational basis for disease classification.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Itoh N, Kayama F, Tatsuki TJ, Tsukamoto T. Have sperm counts deteriorated over the past 20 years in healthy, young Japanese men? Results from the Sapporo area. J. Androl. 2001;22:40–44. - PubMed
Publication types
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
