

Role of intracortical neuropil growth in the gyrification of the primate cerebral cortex
- PMID: 36574666
- PMCID: PMC9910595
- DOI: 10.1073/pnas.2210967120
Role of intracortical neuropil growth in the gyrification of the primate cerebral cortex
Abstract
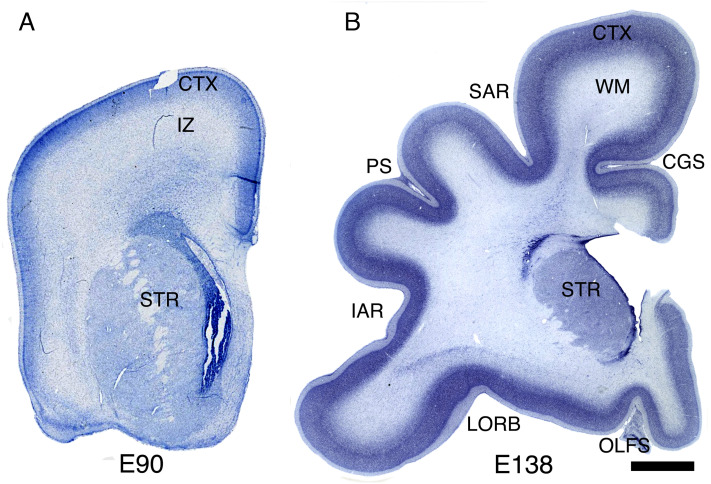
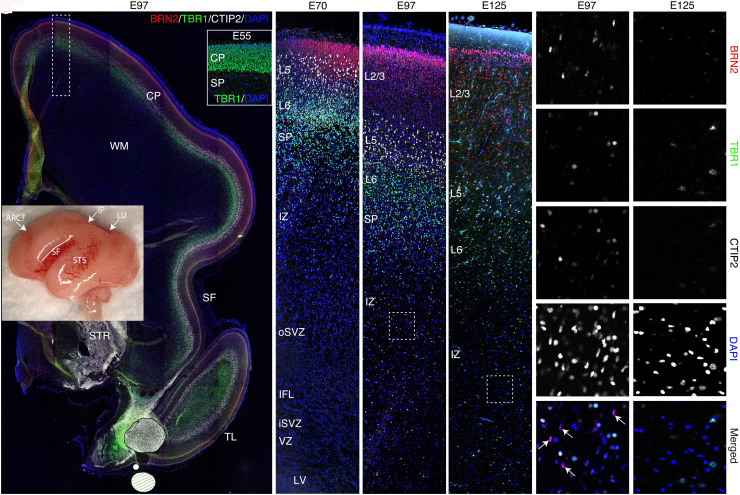
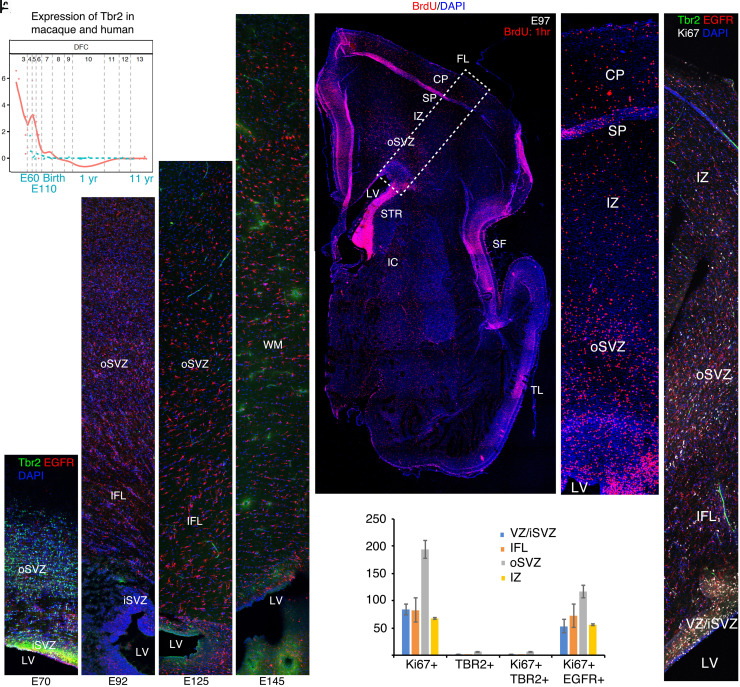
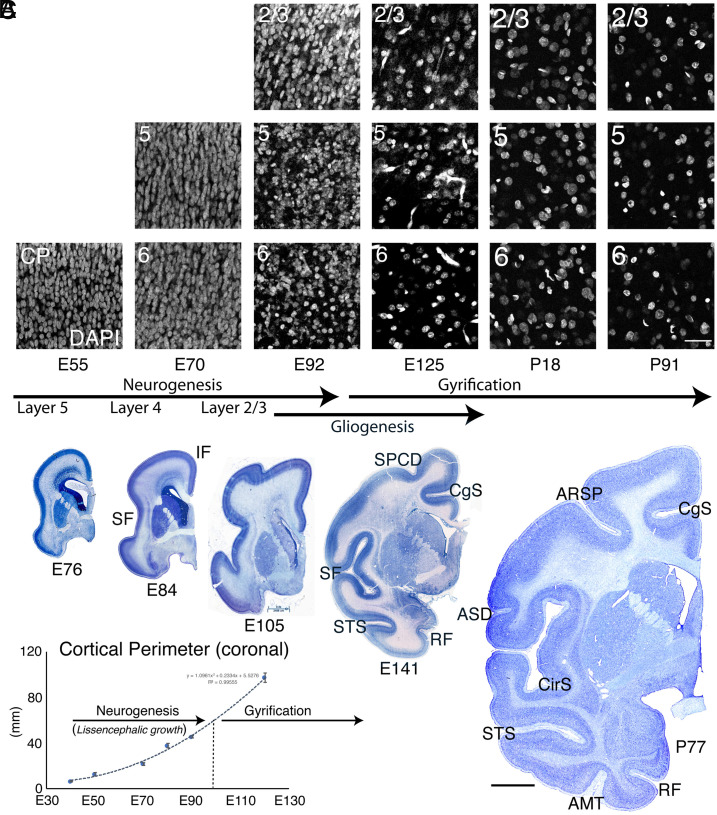
The convolutions of the mammalian cerebral cortex allow the enlargement of its surface and addition of novel functional areas during evolution while minimizing expansion of the cranium. Cognitive neurodevelopmental disorders in humans, including microcephaly and lissencephaly, are often associated with impaired gyrification. In the classical model of gyrification, surface area is initially set by the number of radial units, and the forces driving cortical folding include neuronal growth, formation of neuropil, glial cell intercalation, and the patterned growth of subcortical white matter. An alternative model proposes that specified neurogenic hotspots in the outer subventricular zone (oSVZ) produce larger numbers of neurons that generate convexities in the cortex. This directly contradicts reports showing that cortical neurogenesis and settling of neurons into the cortical plate in primates, including humans, are completed well prior to the formation of secondary and tertiary gyri and indeed most primary gyri. In addition, during the main period of gyrification, the oSVZ produces mainly astrocytes and oligodendrocytes. Here we describe how rapid growth of intracortical neuropil, addition of glial cells, and enlargement of subcortical white matter in primates are the primary forces responsible for the post-neurogenic expansion of the cortical surface and formation of gyri during fetal development. Using immunohistochemistry for markers of proliferation and glial and neuronal progenitors combined with transcriptomic analysis, we show that neurogenesis in the ventricular zone and oSVZ is phased out and transitions to gliogenesis prior to gyral development. In summary, our data support the classical model of gyrification and provide insight into the pathogenesis of congenital cortical malformations.
Keywords: cortex; gyrification; neurogenesis.
Conflict of interest statement
The authors declare no competing interest.
Figures
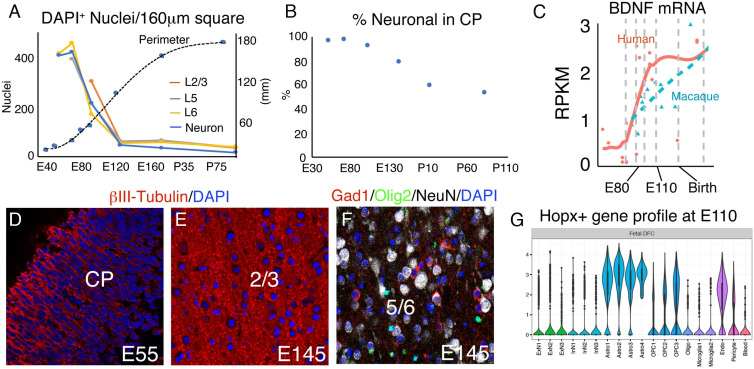
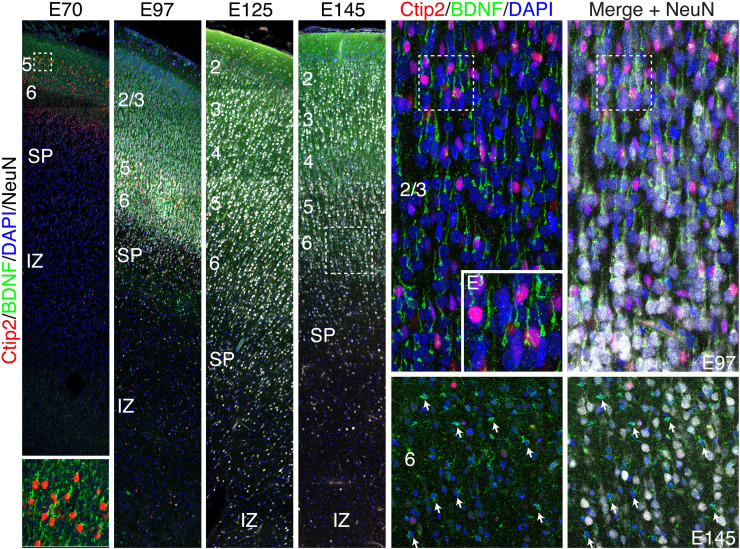
References
-
- Rakic P., Specification of cerebral cortical areas. Science 241, 170–176 (1988). - PubMed
-
- Rakic P., A small step for the cell, a giant leap for mankind: A hypothesis of neocortical expansion during evolution. Trends Neurosci. 18, 383–388 (1995). - PubMed
-
- Dehay C., Kennedy H., Cell-cycle control and cortical development. Nat. Rev. Neurosci. 8, 438–450 (2007). - PubMed
-
- Noctor S. C., Flint A. C., Weissman T. A., Dammerman R. S., Kriegstein A. R., Neurons derived from radial glial cells establish radial units in neocortex. Nature 409, 714–720 (2001). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
