

Obese visceral fat tissue inflammation: from protective to detrimental?
- PMID: 36575472
- PMCID: PMC9795790
- DOI: 10.1186/s12916-022-02672-y
Obese visceral fat tissue inflammation: from protective to detrimental?
Abstract
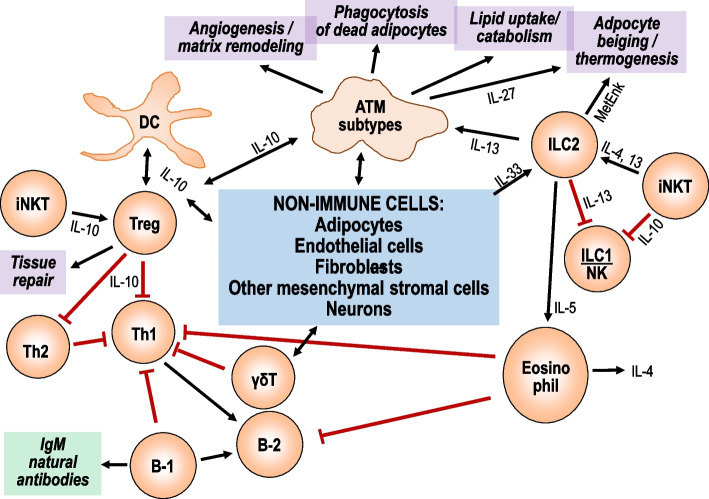
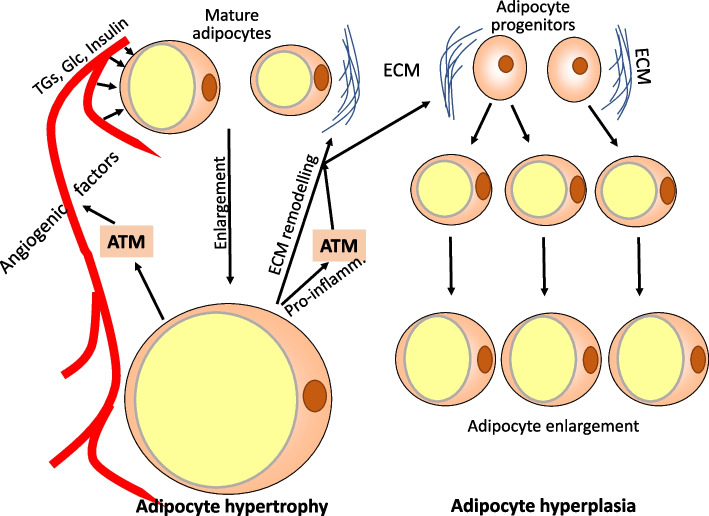
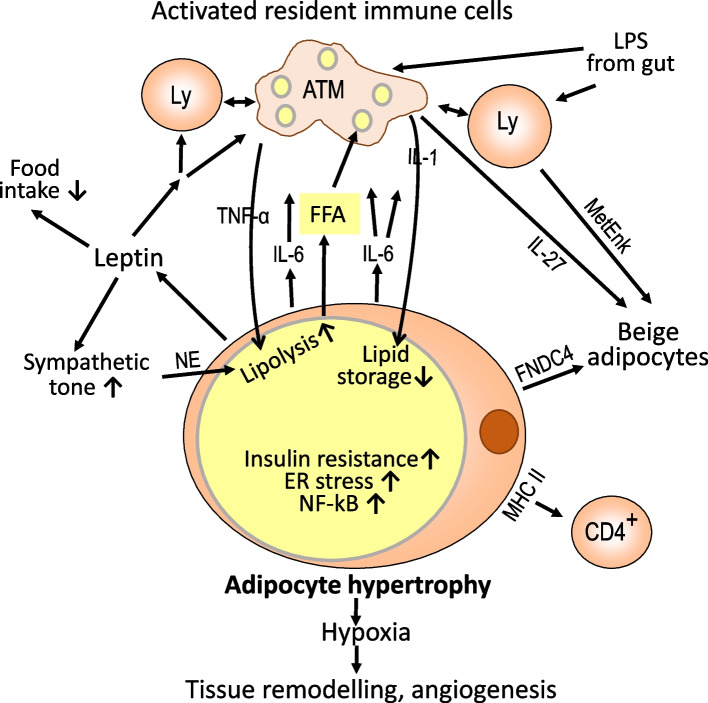
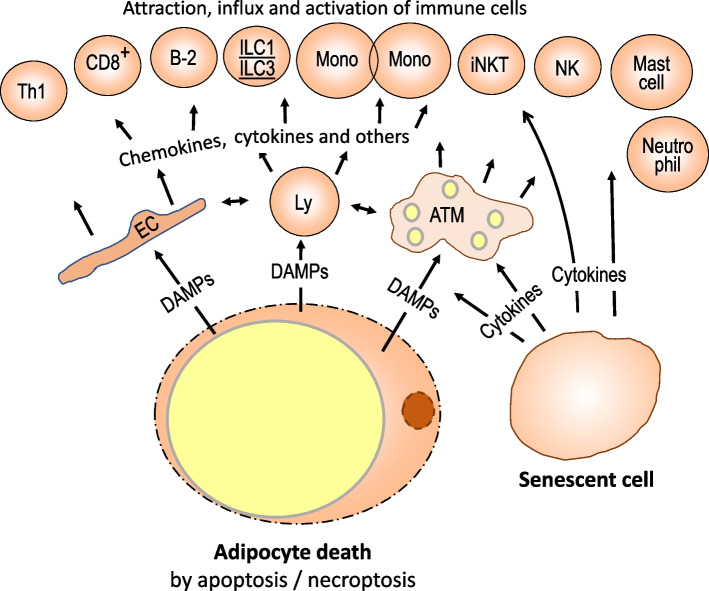
Obesity usually is accompanied by inflammation of fat tissue, with a prominent role of visceral fat. Chronic inflammation in obese fat tissue is of a lower grade than acute immune activation for clearing the tissue from an infectious agent. It is the loss of adipocyte metabolic homeostasis that causes activation of resident immune cells for supporting tissue functions and regaining homeostasis. Initially, the excess influx of lipids and glucose in the context of overnutrition is met by adipocyte growth and proliferation. Eventual lipid overload of hypertrophic adipocytes leads to endoplasmic reticulum stress and the secretion of a variety of signals causing increased sympathetic tone, lipolysis by adipocytes, lipid uptake by macrophages, matrix remodeling, angiogenesis, and immune cell activation. Pro-inflammatory signaling of adipocytes causes the resident immune system to release increased amounts of pro-inflammatory and other mediators resulting in enhanced tissue-protective responses. With chronic overnutrition, these protective actions are insufficient, and death of adipocytes as well as senescence of several tissue cell types is seen. This structural damage causes the expression or release of immunostimulatory cell components resulting in influx and activation of monocytes and many other immune cell types, with a contribution of stromal cells. Matrix remodeling and angiogenesis is further intensified as well as possibly detrimental fibrosis. The accumulation of senescent cells also may be detrimental via eventual spread of senescence state from affected to neighboring cells by the release of microRNA-containing vesicles. Obese visceral fat inflammation can be viewed as an initially protective response in order to cope with excess ambient nutrients and restore tissue homeostasis but may contribute to tissue damage at a later stage.
Keywords: Adipocyte hyperplasia; Adipocyte hypertrophy; Adipose tissue macrophages; Adiposity; Crown-like structures; Cytokines; Inflammation; Obesity; Resident immune cells; Visceral fat.
© 2022. The Author(s).
Conflict of interest statement
The author declares no competing interests.
Figures
References
-
- Weinstock A, Brown EJ, Garabedian ML, Pena S, Sharma M, Lafaille J, et al. Single-Cell RNA Sequencing of Visceral Adipose Tissue Leukocytes Reveals that Caloric Restriction Following Obesity Promotes the Accumulation of a Distinct Macrophage Population with Features of Phagocytic Cells. Immunometabolism. 2019;1:e190008. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
