

Follistatin regulates the specification of the apical cochlea responsible for low-frequency hearing in mammals
- PMID: 36577057
- PMCID: PMC9910458
- DOI: 10.1073/pnas.2213099120
Follistatin regulates the specification of the apical cochlea responsible for low-frequency hearing in mammals
Abstract
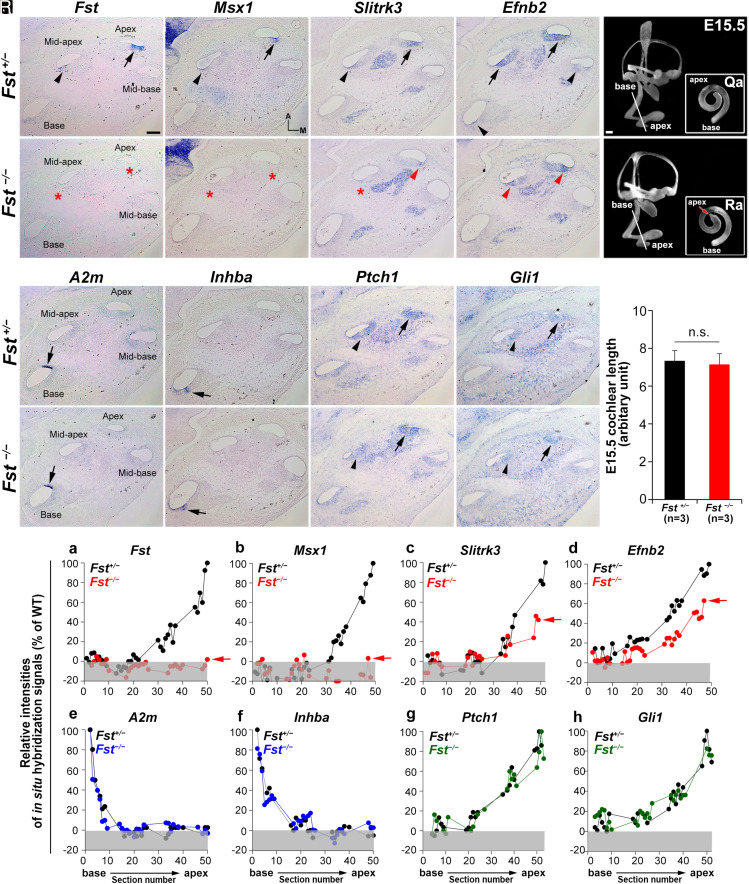
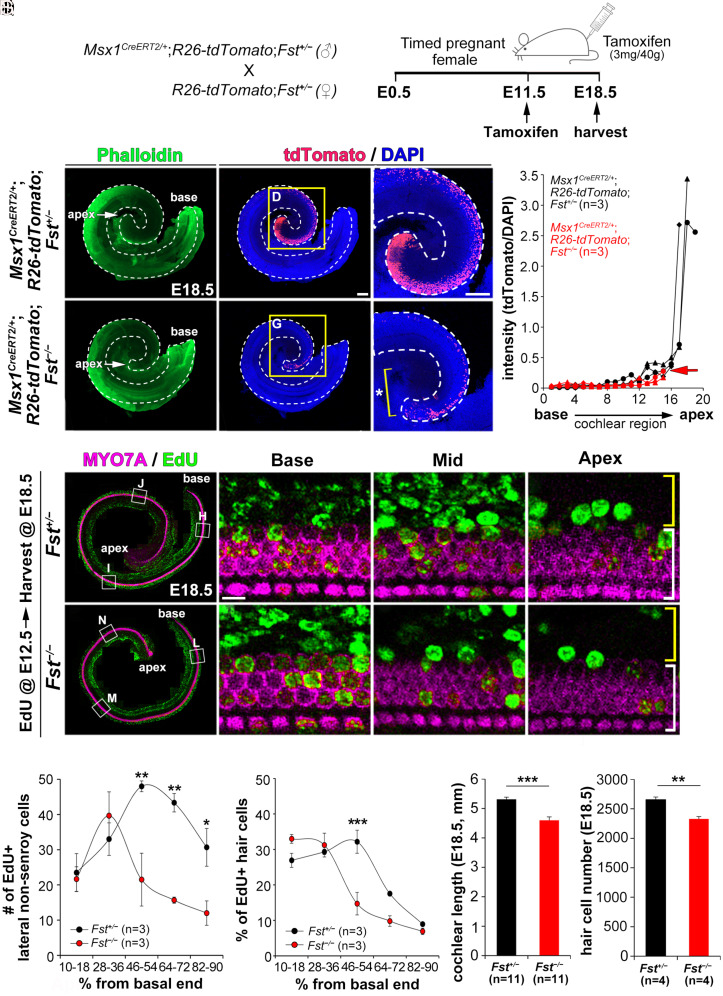
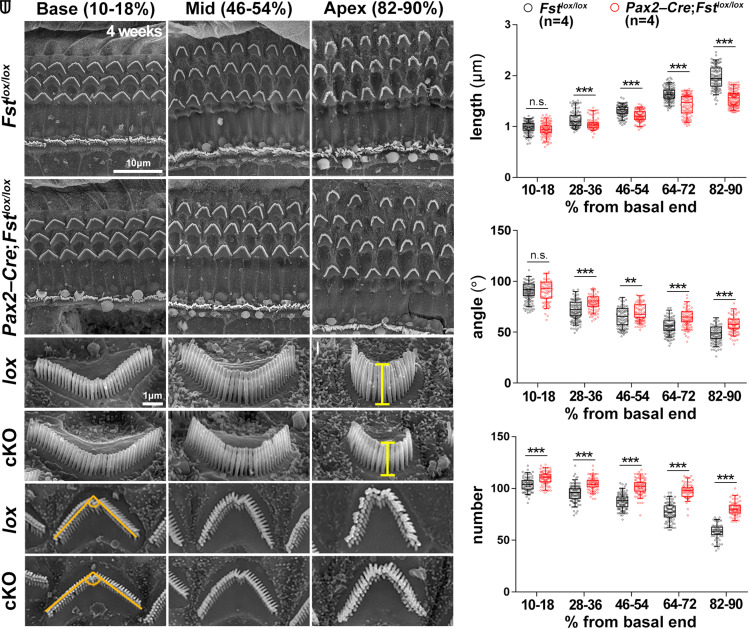
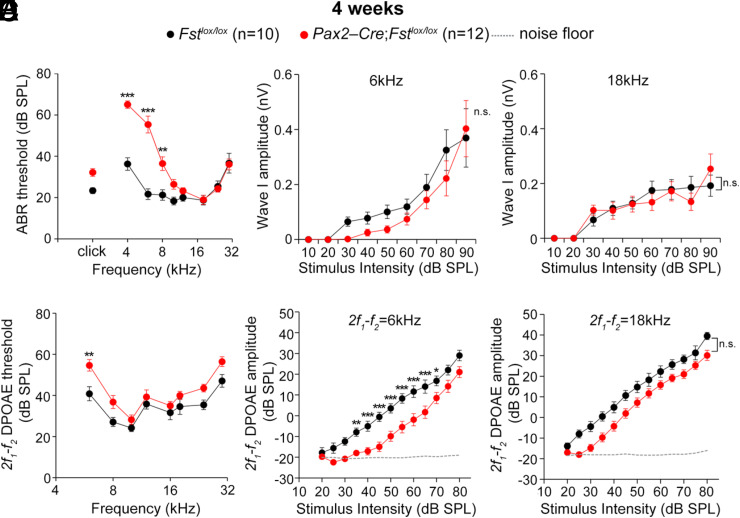
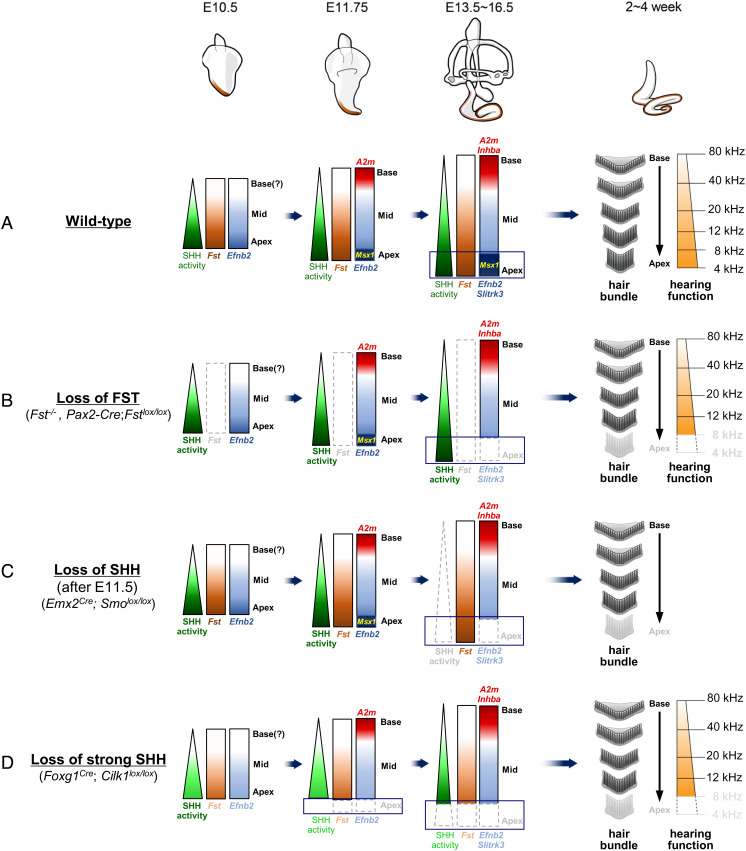
The cochlea's ability to discriminate sound frequencies is facilitated by a special topography along its longitudinal axis known as tonotopy. Auditory hair cells located at the base of the cochlea respond to high-frequency sounds, whereas hair cells at the apex respond to lower frequencies. Gradual changes in morphological and physiological features along the length of the cochlea determine each region's frequency selectivity, but it remains unclear how tonotopy is established during cochlear development. Recently, sonic hedgehog (SHH) was proposed to initiate the establishment of tonotopy by conferring regional identity to the primordial cochlea. Here, using mouse genetics, we provide in vivo evidence that regional identity in the embryonic cochlea acts as a framework upon which tonotopy-specific properties essential for frequency selectivity in the mature cochlea develop. We found that follistatin (FST) is required for the maintenance of apical cochlear identity, but dispensable for its initial induction. In a fate-mapping analysis, we found that FST promotes expansion of apical cochlear cells, contributing to the formation of the apical cochlear domain. SHH, in contrast, is required both for the induction and maintenance of apical identity. In the absence of FST or SHH, mice produce a short cochlea lacking its apical domain. This results in the loss of apex-specific anatomical and molecular properties and low-frequency-specific hearing loss.
Keywords: cochlea; follistatin; frequency discrimination; tonotopy.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Conserved role of Sonic Hedgehog in tonotopic organization of the avian basilar papilla and mammalian cochlea.Proc Natl Acad Sci U S A. 2015 Mar 24;112(12):3746-51. doi: 10.1073/pnas.1417856112. Epub 2015 Mar 9. Proc Natl Acad Sci U S A. 2015. PMID: 25775517 Free PMC article.
-
Developmental gene expression profiling along the tonotopic axis of the mouse cochlea.PLoS One. 2012;7(7):e40735. doi: 10.1371/journal.pone.0040735. Epub 2012 Jul 12. PLoS One. 2012. PMID: 22808246 Free PMC article.
-
Place Coding in the Human Cochlea.medRxiv [Preprint]. 2023 May 22:2023.04.13.23288518. doi: 10.1101/2023.04.13.23288518. medRxiv. 2023. Update in: Ear Hear. 2025 Jan-Feb 01;46(1):253-264. doi: 10.1097/AUD.0000000000001579. PMID: 37131618 Free PMC article. Updated. Preprint.
-
Where hearing starts: the development of the mammalian cochlea.J Anat. 2016 Feb;228(2):233-54. doi: 10.1111/joa.12314. Epub 2015 Jun 5. J Anat. 2016. PMID: 26052920 Free PMC article. Review.
-
Signatures of cochlear processing in neuronal coding of auditory information.Mol Cell Neurosci. 2022 May;120:103732. doi: 10.1016/j.mcn.2022.103732. Epub 2022 Apr 27. Mol Cell Neurosci. 2022. PMID: 35489636 Review.
Cited by
-
MYH1 deficiency disrupts outer hair cell electromotility, resulting in hearing loss.Exp Mol Med. 2024 Nov;56(11):2423-2435. doi: 10.1038/s12276-024-01338-4. Epub 2024 Nov 1. Exp Mol Med. 2024. PMID: 39482536 Free PMC article.
-
Early Steps towards Hearing: Placodes and Sensory Development.Int J Mol Sci. 2023 Apr 10;24(8):6994. doi: 10.3390/ijms24086994. Int J Mol Sci. 2023. PMID: 37108158 Free PMC article. Review.
-
3D reconstruction of the mouse cochlea from scRNA-seq data suggests morphogen-based principles in apex-to-base specification.Dev Cell. 2024 Jun 17;59(12):1538-1552.e6. doi: 10.1016/j.devcel.2024.03.028. Epub 2024 Apr 8. Dev Cell. 2024. PMID: 38593801 Free PMC article.
-
Alternative splicing in shaping the molecular landscape of the cochlea.Front Cell Dev Biol. 2023 Mar 2;11:1143428. doi: 10.3389/fcell.2023.1143428. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 36936679 Free PMC article. Review.
-
SHANK2 establishes auditory hair bundle architecture essential for mammalian hearing.Proc Natl Acad Sci U S A. 2025 Jul 15;122(28):e2426646122. doi: 10.1073/pnas.2426646122. Epub 2025 Jul 8. Proc Natl Acad Sci U S A. 2025. PMID: 40627398
References
-
- Mann Z. F., Kelley M. W., Development of tonotopy in the auditory periphery. Hear. Res. 276, 2–15 (2011). - PubMed
-
- Davis R. L., Gradients of neurotrophins, ion channels, and tuning in the cochlea. Neuroscientist 9, 311–316 (2003). - PubMed
-
- Wright A., Dimensions of the cochlear stereocilia in man and the guinea pig. Hear. Res. 13, 89–98 (1984). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
