

Bone marrow-derived IGF-1 orchestrates maintenance and regeneration of the adult skeleton
- PMID: 36577075
- PMCID: PMC9910602
- DOI: 10.1073/pnas.2203779120
Bone marrow-derived IGF-1 orchestrates maintenance and regeneration of the adult skeleton
Abstract
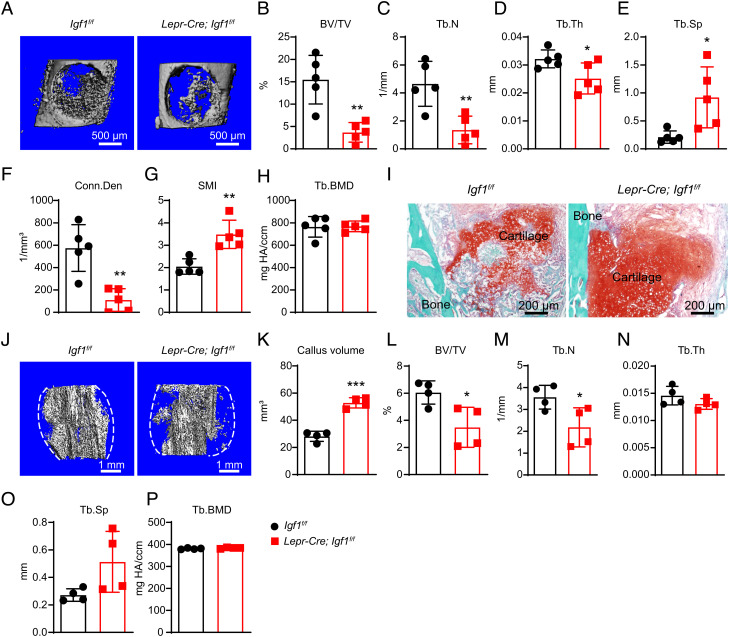
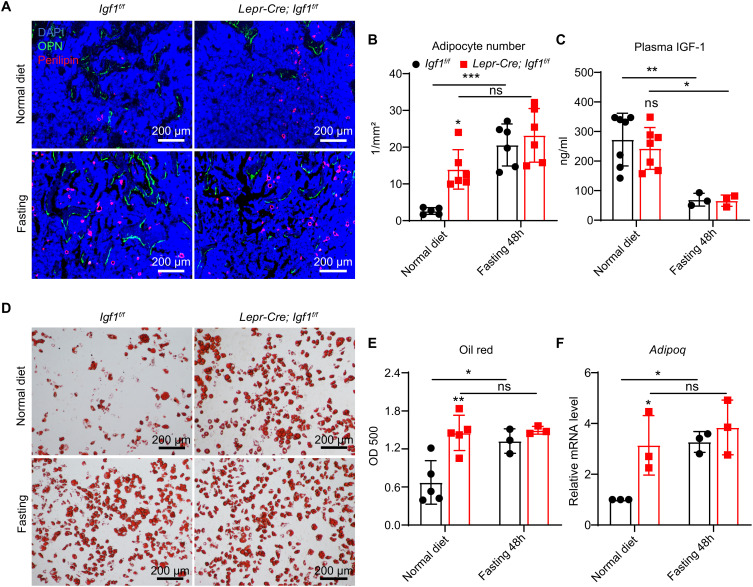
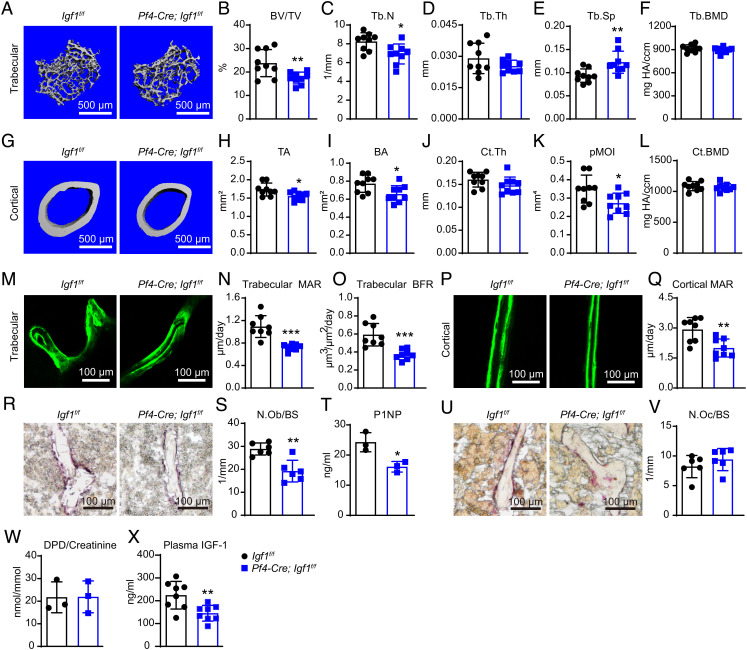
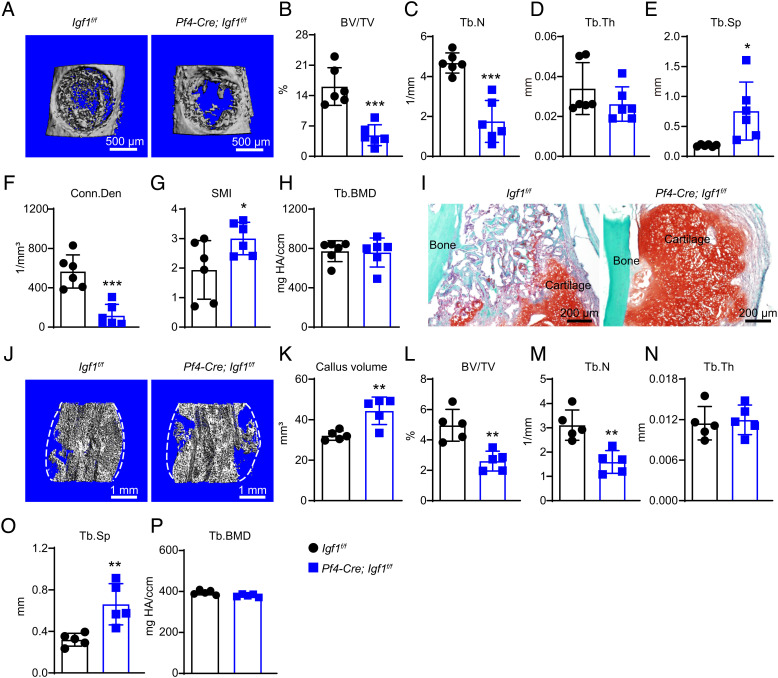
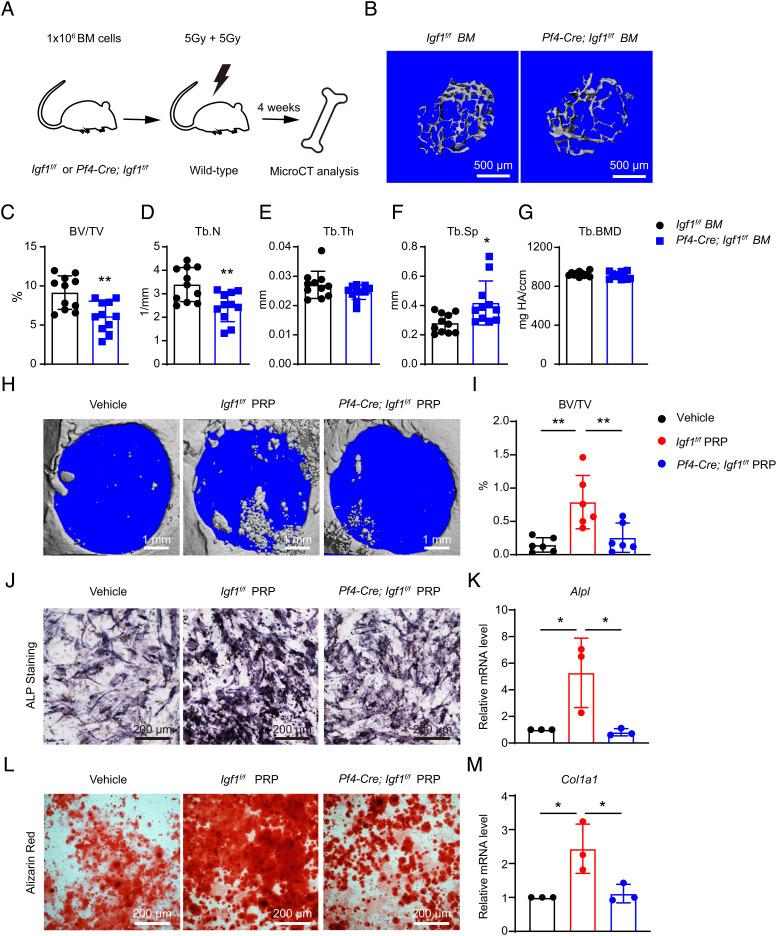
Insulin-like growth factor I (IGF-1) is a key regulator of tissue growth and development in response to growth hormone stimulation. In the skeletal system, IGF-1 derived from osteoblasts and chondrocytes are essential for normal bone development; however, whether bone marrow (BM)-resident cells provide distinct sources of IGF-1 in the adult skeleton remains elusive. Here, we show that BM stromal cells (BMSCs) and megakaryocytes/platelets (MKs/PLTs) express the highest levels of IGF-1 in adult long bones. Deletion of Igf1 from BMSCs by Lepr-Cre leads to decreased bone formation, impaired bone regeneration, and increased BM adipogenesis. Importantly, reduction of BMSC-derived IGF-1 contributes to fasting-induced marrow fat accumulation. In contrast, deletion of Igf1 from MKs/PLTs by Pf4-Cre leads to reduced bone formation and regeneration without affecting BM adipogenesis. To our surprise, MKs/PLTs are also an important source of systemic IGF-1. Platelet-rich plasma (PRP) from Pf4-Cre; Igf1f/fmice showed compromised osteogenic potential both in vivo and in vitro, suggesting that MK/PLT-derived IGF-1 underlies the therapeutic effects of PRP. Taken together, this study identifies BMSCs and MKs/PLTs as two important sources of IGF-1 that coordinate to maintain and regenerate the adult skeleton, highlighting reciprocal regulation between the hematopoietic and skeletal systems.
Keywords: IGF-1; adipogenesis; bone marrow; hematopoiesis; osteogenesis.
Conflict of interest statement
The authors declare no competing interest.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
