

Disruption of the ATXN1-CIC complex reveals the role of additional nuclear ATXN1 interactors in spinocerebellar ataxia type 1
- PMID: 36577402
- PMCID: PMC9957872
- DOI: 10.1016/j.neuron.2022.11.016
Disruption of the ATXN1-CIC complex reveals the role of additional nuclear ATXN1 interactors in spinocerebellar ataxia type 1
Erratum in
-
Disruption of the ATXN1-CIC complex reveals the role of additional nuclear ATXN1 interactors in spinocerebellar ataxia type 1.Neuron. 2023 Mar 15;111(6):915. doi: 10.1016/j.neuron.2023.02.030. Neuron. 2023. PMID: 36924764 Free PMC article. No abstract available.
Abstract
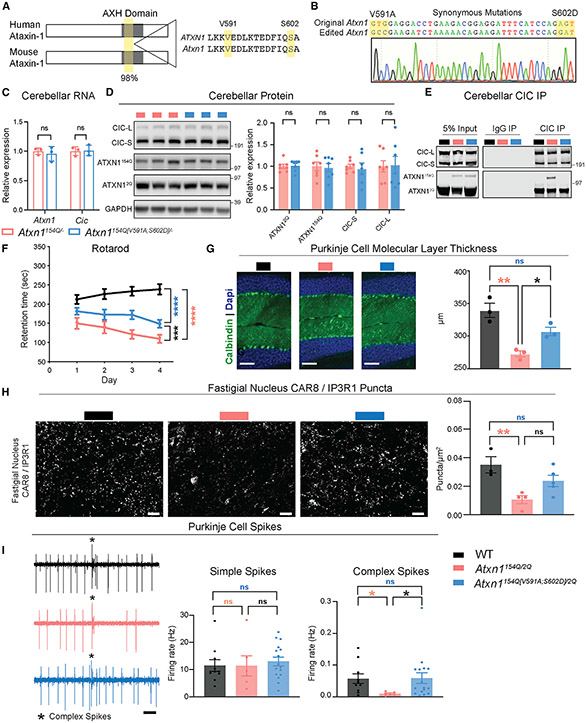
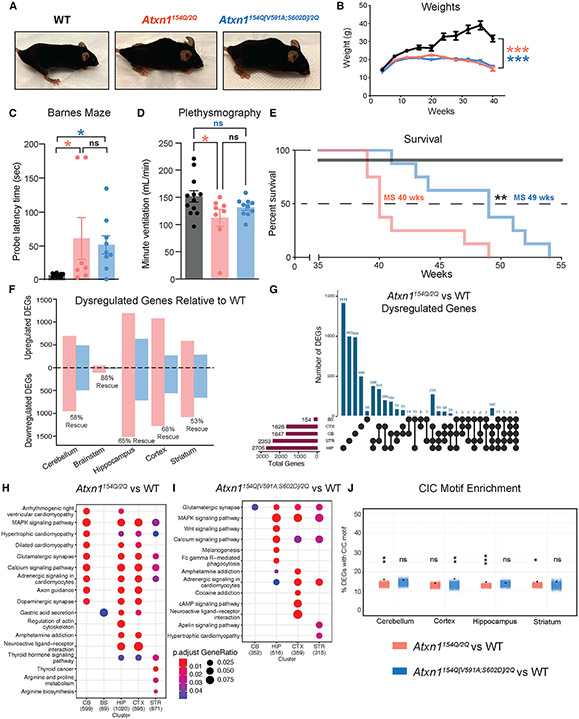
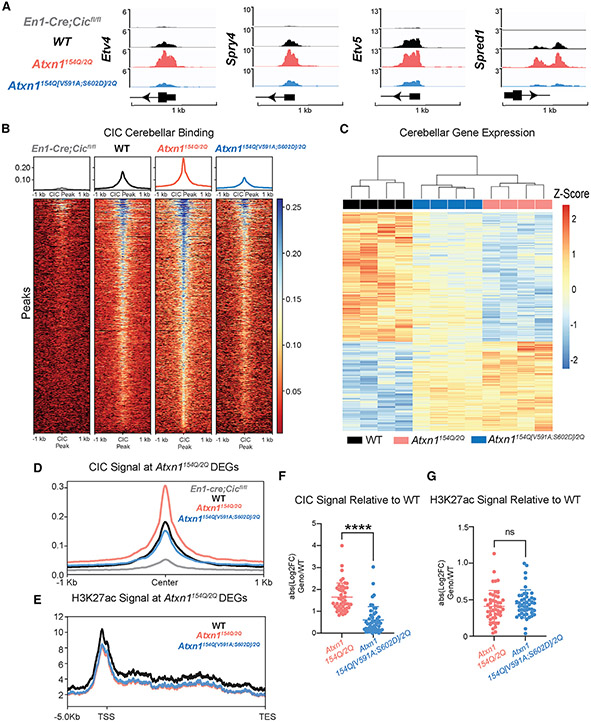
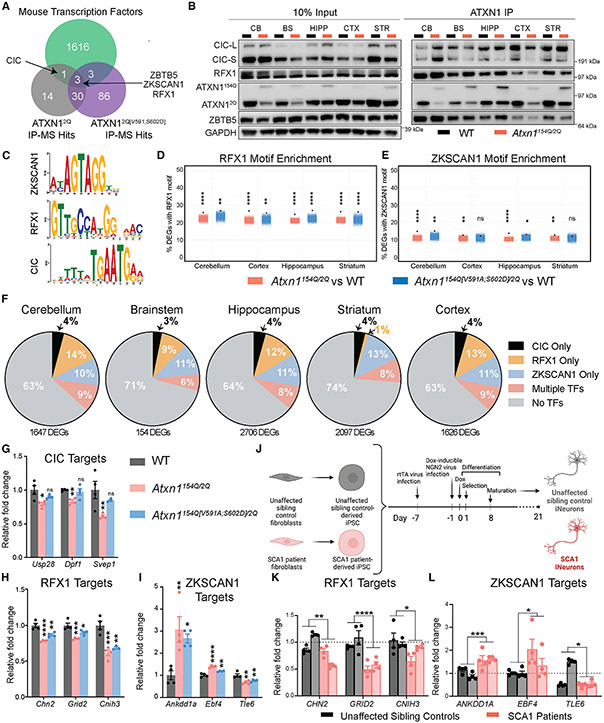
Spinocerebellar ataxia type 1 (SCA1) is a paradigmatic neurodegenerative disease in that it is caused by a mutation in a broadly expressed protein, ATXN1; however, only select populations of cells degenerate. The interaction of polyglutamine-expanded ATXN1 with the transcriptional repressor CIC drives cerebellar Purkinje cell pathogenesis; however, the importance of this interaction in other vulnerable cells remains unknown. Here, we mutated the 154Q knockin allele of Atxn1154Q/2Q mice to prevent the ATXN1-CIC interaction globally. This normalized genome-wide CIC binding; however, it only partially corrected transcriptional and behavioral phenotypes, suggesting the involvement of additional factors in disease pathogenesis. Using unbiased proteomics, we identified three ATXN1-interacting transcription factors: RFX1, ZBTB5, and ZKSCAN1. We observed altered expression of RFX1 and ZKSCAN1 target genes in SCA1 mice and patient-derived iNeurons, highlighting their potential contributions to disease. Together, these data underscore the complexity of mechanisms driving cellular vulnerability in SCA1.
Keywords: ATXN1; CIC; CUT&RUN; SCA1; neurodegeneration; polyglutamine expansion disorders; proteomics; regional vulnerability; spinocerebellar ataxia type 1; transcriptomics.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests H.Y.Z. is a co-founder of Cajal Neuroscience and a member of its scientific advisory board. H.Y.Z. is a science partner at the Column Group and a board member of Regeneron. H.Y.Z. collaborates with UCB and Ionis Pharmaceuticals on projects not relevant to this publication.
Figures
Comment in
-
Decreasing mutant ATXN1 nuclear localization improves a spectrum of SCA1-like phenotypes and brain region transcriptomic profiles.Neuron. 2023 Feb 15;111(4):493-507.e6. doi: 10.1016/j.neuron.2022.11.017. Epub 2022 Dec 27. Neuron. 2023. PMID: 36577403 Free PMC article.
-
Diverse regional mechanisms drive spinocerebellar ataxia type 1 phenotypes.Neuron. 2023 Feb 15;111(4):447-449. doi: 10.1016/j.neuron.2023.01.021. Neuron. 2023. PMID: 36796325
References
-
- Opal P, and Ashizawa T (1998). Spinocerebellar ataxia type 1. GeneReviews (University of Washington; ), pp. 1–12. https://www.ncbi.nlm.nih.gov/books/NBK1184/.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
