

High-efficiency production of 5-hydroxyectoine using metabolically engineered Corynebacterium glutamicum
- PMID: 36578077
- PMCID: PMC9798599
- DOI: 10.1186/s12934-022-02003-z
High-efficiency production of 5-hydroxyectoine using metabolically engineered Corynebacterium glutamicum
Abstract
Background: Extremolytes enable microbes to withstand even the most extreme conditions in nature. Due to their unique protective properties, the small organic molecules, more and more, become high-value active ingredients for the cosmetics and the pharmaceutical industries. While ectoine, the industrial extremolyte flagship, has been successfully commercialized before, an economically viable route to its highly interesting derivative 5-hydroxyectoine (hydroxyectoine) is not existing.
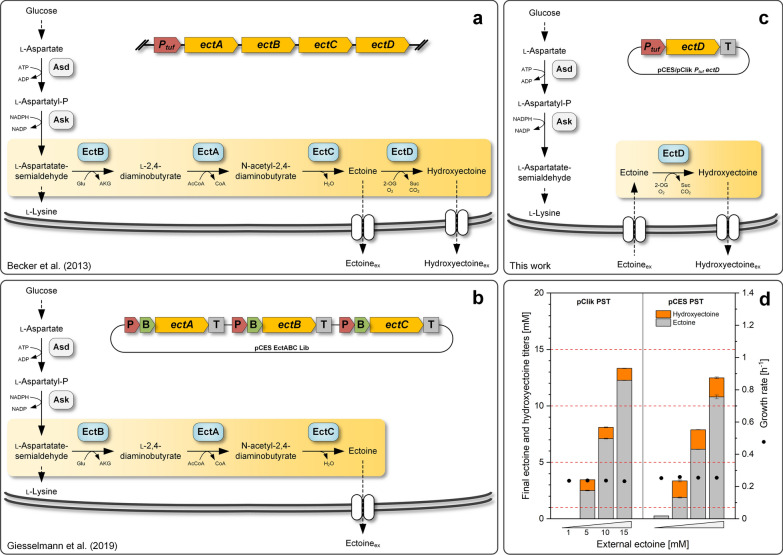
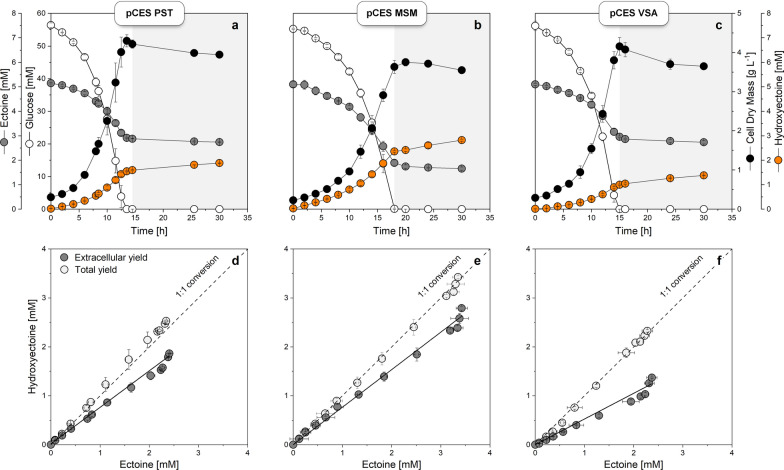
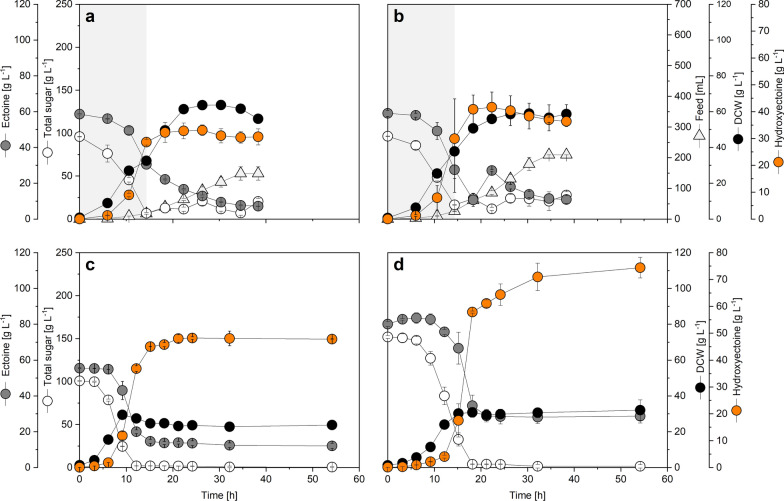
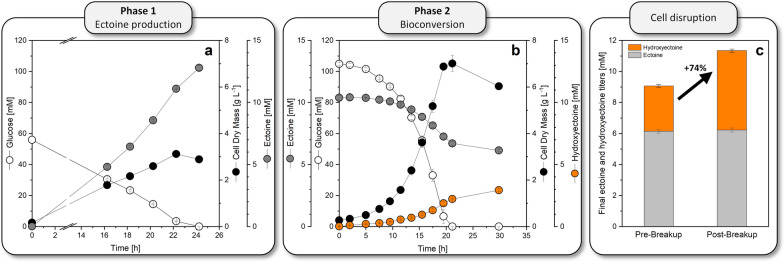
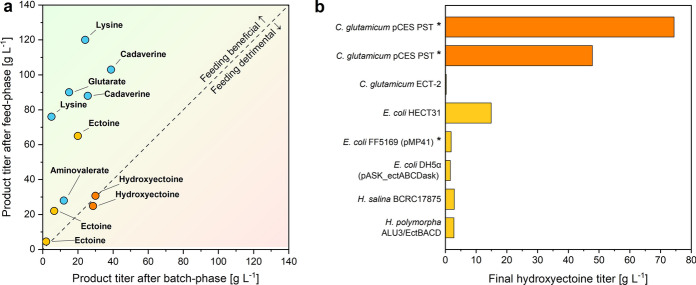
Results: Here, we demonstrate high-level hydroxyectoine production, using metabolically engineered strains of C. glutamicum that express a codon-optimized, heterologous ectD gene, encoding for ectoine hydroxylase, to convert supplemented ectoine in the presence of sucrose as growth substrate into the desired derivative. Fourteen out of sixteen codon-optimized ectD variants from phylogenetically diverse bacterial and archaeal donors enabled hydroxyectoine production, showing the strategy to work almost regardless of the origin of the gene. The genes from Pseudomonas stutzeri (PST) and Mycobacterium smegmatis (MSM) worked best and enabled hydroxyectoine production up to 97% yield. Metabolic analyses revealed high enrichment of the ectoines inside the cells, which, inter alia, reduced the synthesis of other compatible solutes, including proline and trehalose. After further optimization, C. glutamicum Ptuf ectDPST achieved a titre of 74 g L-1 hydroxyectoine at 70% selectivity within 12 h, using a simple batch process. In a two-step procedure, hydroxyectoine production from ectoine, previously synthesized fermentatively with C. glutamicum ectABCopt, was successfully achieved without intermediate purification.
Conclusions: C. glutamicum is a well-known and industrially proven host, allowing the synthesis of commercial products with granted GRAS status, a great benefit for a safe production of hydroxyectoine as active ingredient for cosmetic and pharmaceutical applications. Because ectoine is already available at commercial scale, its use as precursor appears straightforward. In the future, two-step processes might provide hydroxyectoine de novo from sugar.
Keywords: 5-Hydroxyectoine; Biotransformation; Corynebacterium glutamicum; Ectoine; Ectoine hydroxylase; Extremolyte; High-value product; Intracellular metabolite; Proline; Trehalose.
© 2022. The Author(s).
Conflict of interest statement
Lukas Jungmann, Sarah Lisa Hoffmann, Caroline Lang, and Christoph Wittmann are co-inventors on a patent application related to this work. The other authors declare no competing interests.
Figures


Similar articles
-
EctD-mediated biotransformation of the chemical chaperone ectoine into hydroxyectoine and its mechanosensitive channel-independent excretion.Microb Cell Fact. 2016 Jul 20;15(1):126. doi: 10.1186/s12934-016-0525-4. Microb Cell Fact. 2016. PMID: 27439307 Free PMC article.
-
Metabolic Engineering of Corynebacterium glutamicum for High-Level Ectoine Production: Design, Combinatorial Assembly, and Implementation of a Transcriptionally Balanced Heterologous Ectoine Pathway.Biotechnol J. 2019 Sep;14(9):e1800417. doi: 10.1002/biot.201800417. Epub 2019 Jul 3. Biotechnol J. 2019. PMID: 31106985
-
The ectD gene, which is involved in the synthesis of the compatible solute hydroxyectoine, is essential for thermoprotection of the halophilic bacterium Chromohalobacter salexigens.J Bacteriol. 2006 Jun;188(11):3774-84. doi: 10.1128/JB.00136-06. J Bacteriol. 2006. PMID: 16707670 Free PMC article.
-
Microbial production of ectoine and hydroxyectoine as high-value chemicals.Microb Cell Fact. 2021 Mar 26;20(1):76. doi: 10.1186/s12934-021-01567-6. Microb Cell Fact. 2021. PMID: 33771157 Free PMC article. Review.
-
Multiple Functions of Compatible Solute Ectoine and Strategies for Constructing Overproducers for Biobased Production.Mol Biotechnol. 2024 Aug;66(8):1772-1785. doi: 10.1007/s12033-023-00827-7. Epub 2023 Jul 24. Mol Biotechnol. 2024. PMID: 37488320 Review.
Cited by
-
Elucidating the salt-tolerant mechanism of Halomonas cupida J9 and unsterile ectoine production from lignocellulosic biomass.Microb Cell Fact. 2024 Aug 31;23(1):237. doi: 10.1186/s12934-024-02515-w. Microb Cell Fact. 2024. PMID: 39217338 Free PMC article.
-
Rational engineering of Halomonas salifodinae to enhance hydroxyectoine production under lower-salt conditions.Appl Microbiol Biotechnol. 2024 May 31;108(1):353. doi: 10.1007/s00253-024-13197-0. Appl Microbiol Biotechnol. 2024. PMID: 38819481 Free PMC article.
-
Metabolic Profile of the Genome-Reduced Bacillus subtilis Strain IIG-Bs-27-39: An Attractive Chassis for Recombinant Protein Production.ACS Synth Biol. 2024 Jul 19;13(7):2199-2214. doi: 10.1021/acssynbio.4c00254. Epub 2024 Jul 9. ACS Synth Biol. 2024. PMID: 38981062 Free PMC article.
-
Complete genome sequence of a haloalkaliphilic heterotrophic bacterium Halomonas salifodinae IM328.Microbiol Resour Announc. 2024 Jul 18;13(7):e0008424. doi: 10.1128/mra.00084-24. Epub 2024 Jun 11. Microbiol Resour Announc. 2024. PMID: 38860781 Free PMC article.
-
Unlocking Ectoine's Postbiotic Therapeutic Promise: Mechanisms, Applications, and Future Directions.Probiotics Antimicrob Proteins. 2025 Mar 12. doi: 10.1007/s12602-025-10506-5. Online ahead of print. Probiotics Antimicrob Proteins. 2025. PMID: 40072821 Review.
References
-
- Schwibbert K, Marin-Sanguino A, Bagyan I, Heidrich G, Lentzen G, Seitz H, Rampp M, Schuster SC, Klenk H-P, Pfeiffer F, et al. A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581 T. Environ Microbiol. 2011;13:1973–1994. doi: 10.1111/j.1462-2920.2010.02336.x. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- EXTRA (031B0822A)/Bundesministerium für Bildung und Forschung
- EXTRA (031B0822A)/Bundesministerium für Bildung und Forschung
- EXTRA (031B0822A)/Bundesministerium für Bildung und Forschung
- EXTRA (031B0822A)/Bundesministerium für Bildung und Forschung
- EXTRA (031B0822A)/Bundesministerium für Bildung und Forschung
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
