

Aortic Stress Activates an Adaptive Program in Thoracic Aortic Smooth Muscle Cells That Maintains Aortic Strength and Protects Against Aneurysm and Dissection in Mice
- PMID: 36579645
- PMCID: PMC9877188
- DOI: 10.1161/ATVBAHA.122.318135
Aortic Stress Activates an Adaptive Program in Thoracic Aortic Smooth Muscle Cells That Maintains Aortic Strength and Protects Against Aneurysm and Dissection in Mice
Abstract
Background: When aortic cells are under stress, such as increased hemodynamic pressure, they adapt to the environment by modifying their functions, allowing the aorta to maintain its strength. To understand the regulation of this adaptive response, we examined transcriptomic and epigenomic programs in aortic smooth muscle cells (SMCs) during the adaptive response to AngII (angiotensin II) infusion and determined its importance in protecting against aortic aneurysm and dissection (AAD).
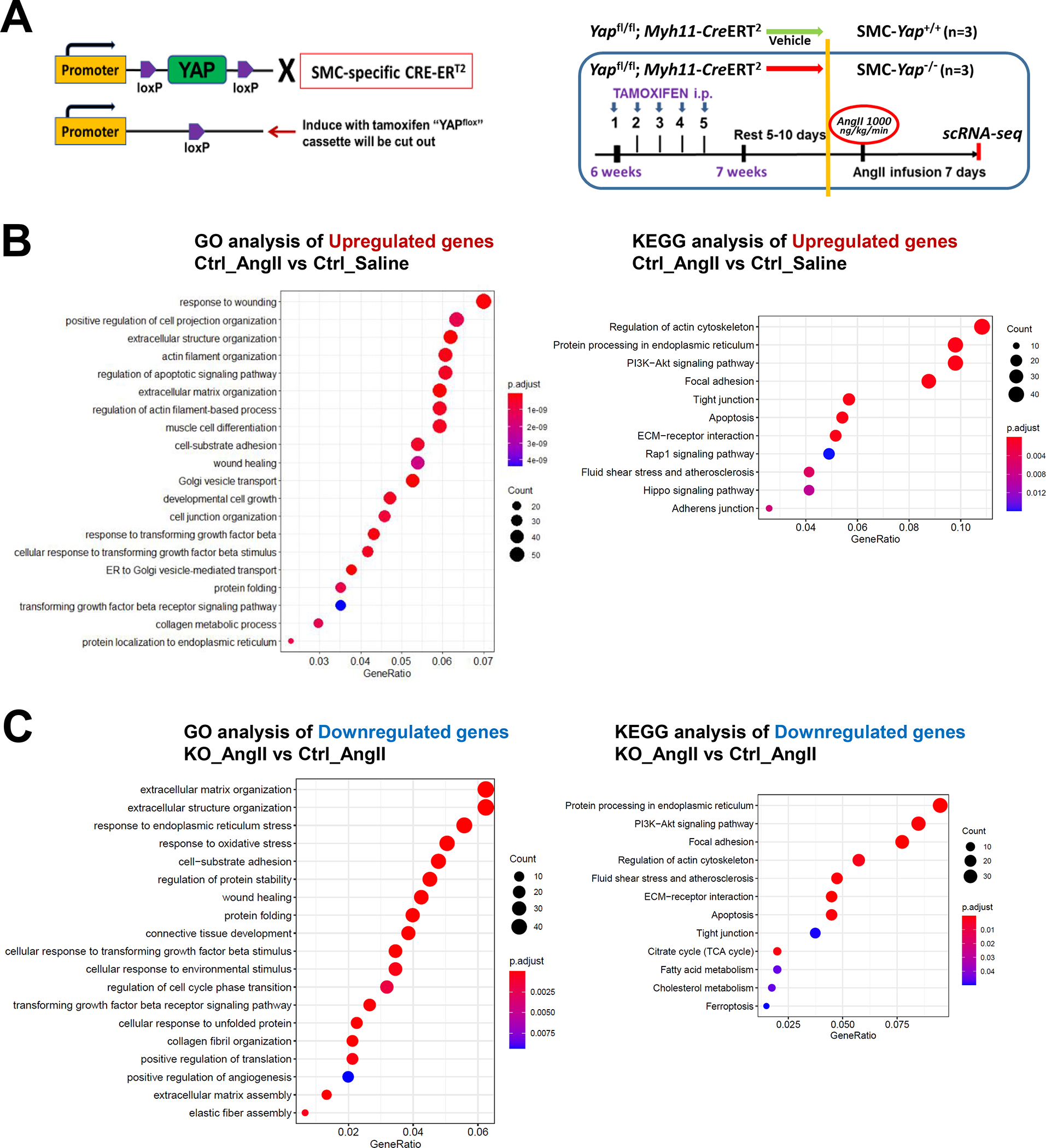
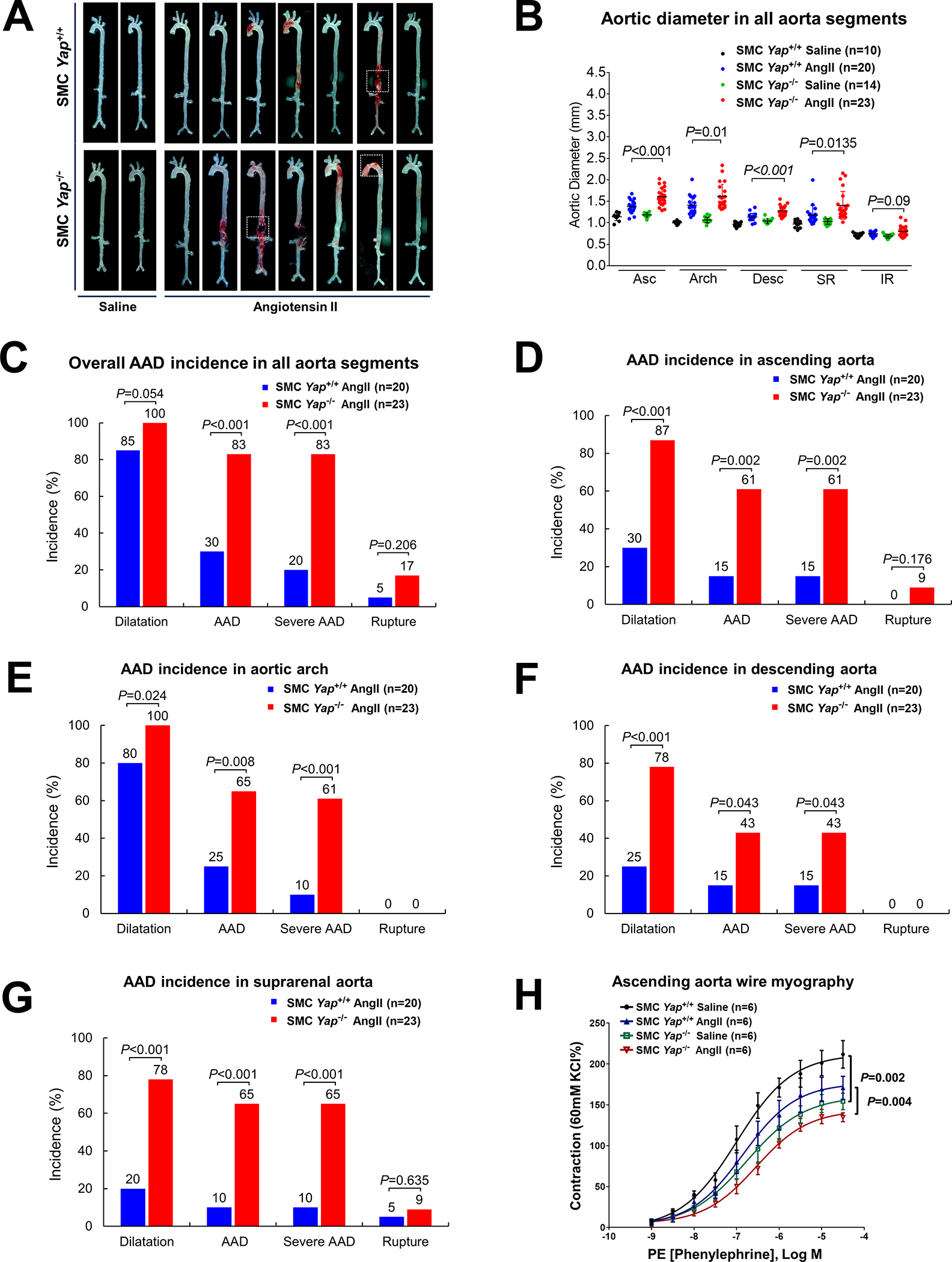
Methods: We performed single-cell RNA sequencing and single-cell sequencing assay for transposase-accessible chromatin (scATAC-seq) analyses in a mouse model of sporadic AAD induced by AngII infusion. We also examined the direct effects of YAP (yes-associated protein) on the SMC adaptive response in vitro. The role of YAP in AAD development was further evaluated in AngII-infused mice with SMC-specific Yap deletion.
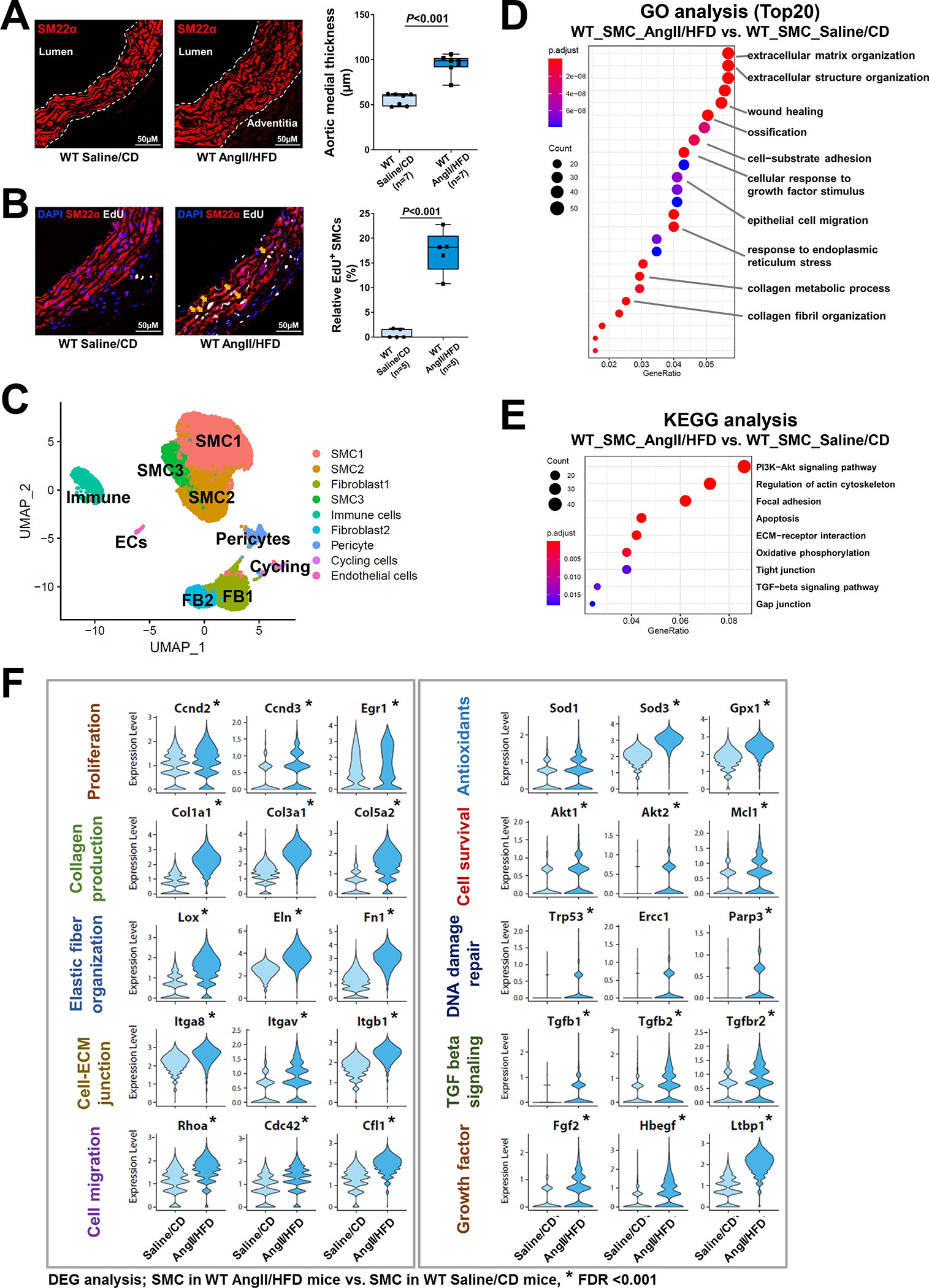
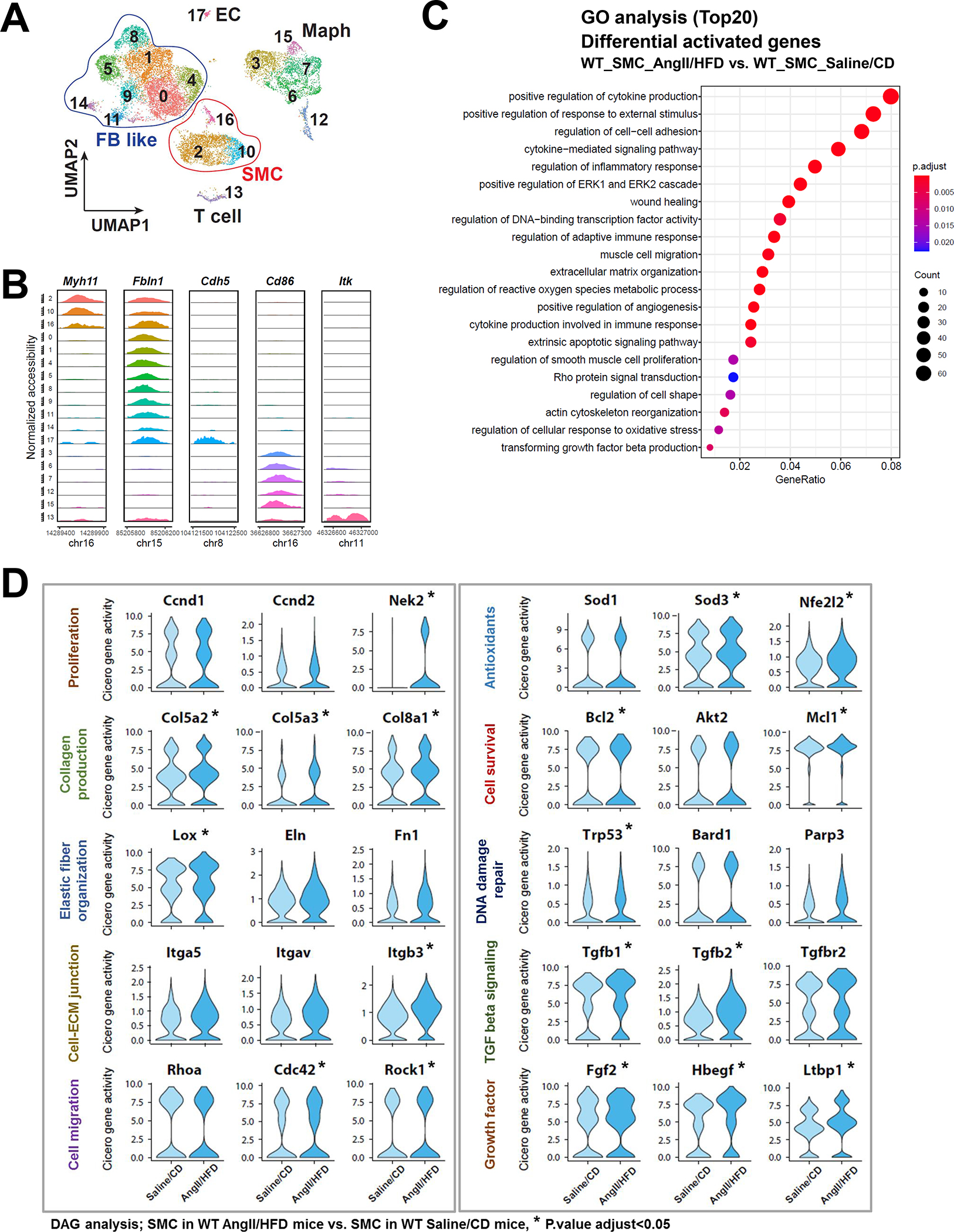
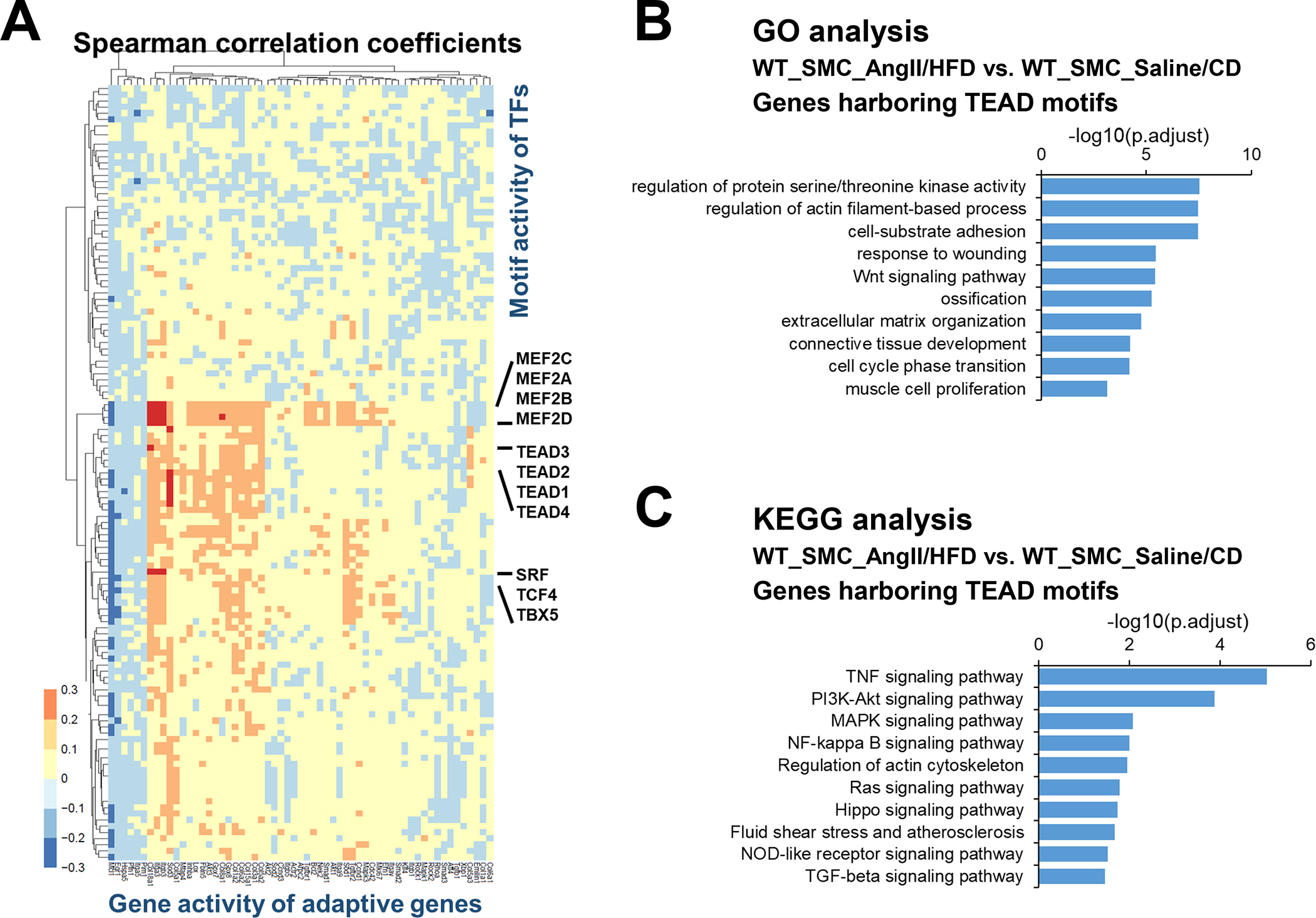
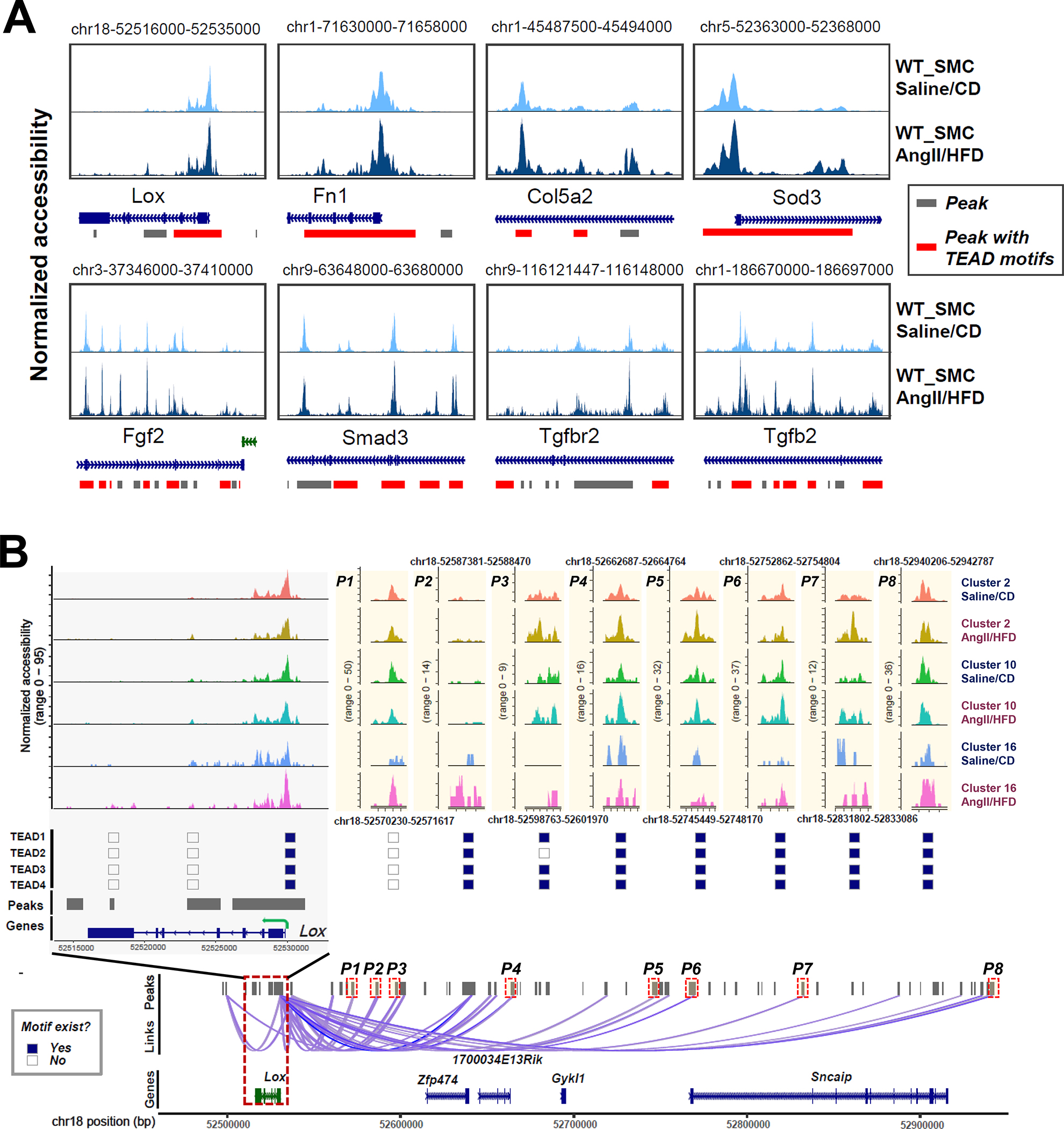
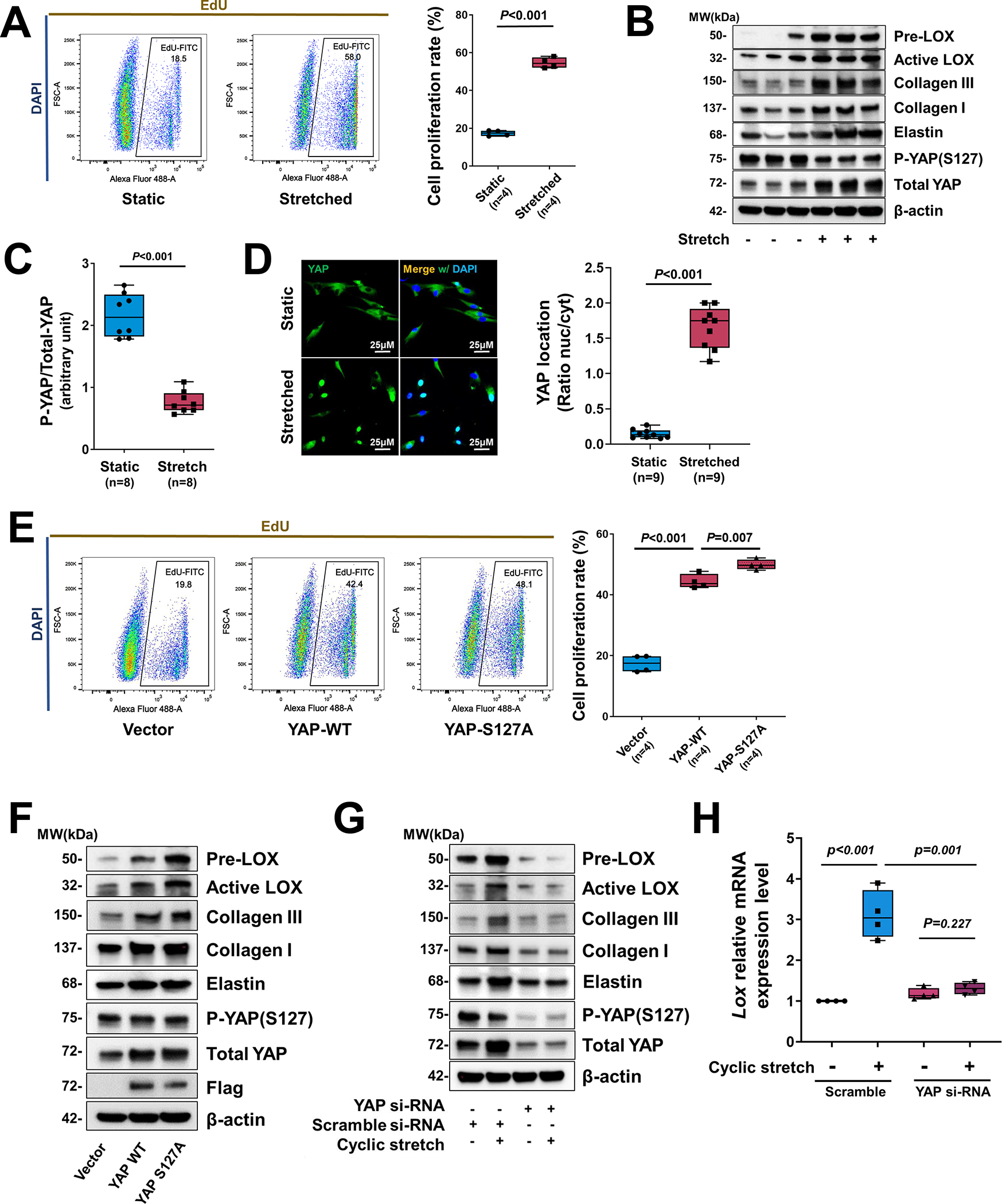
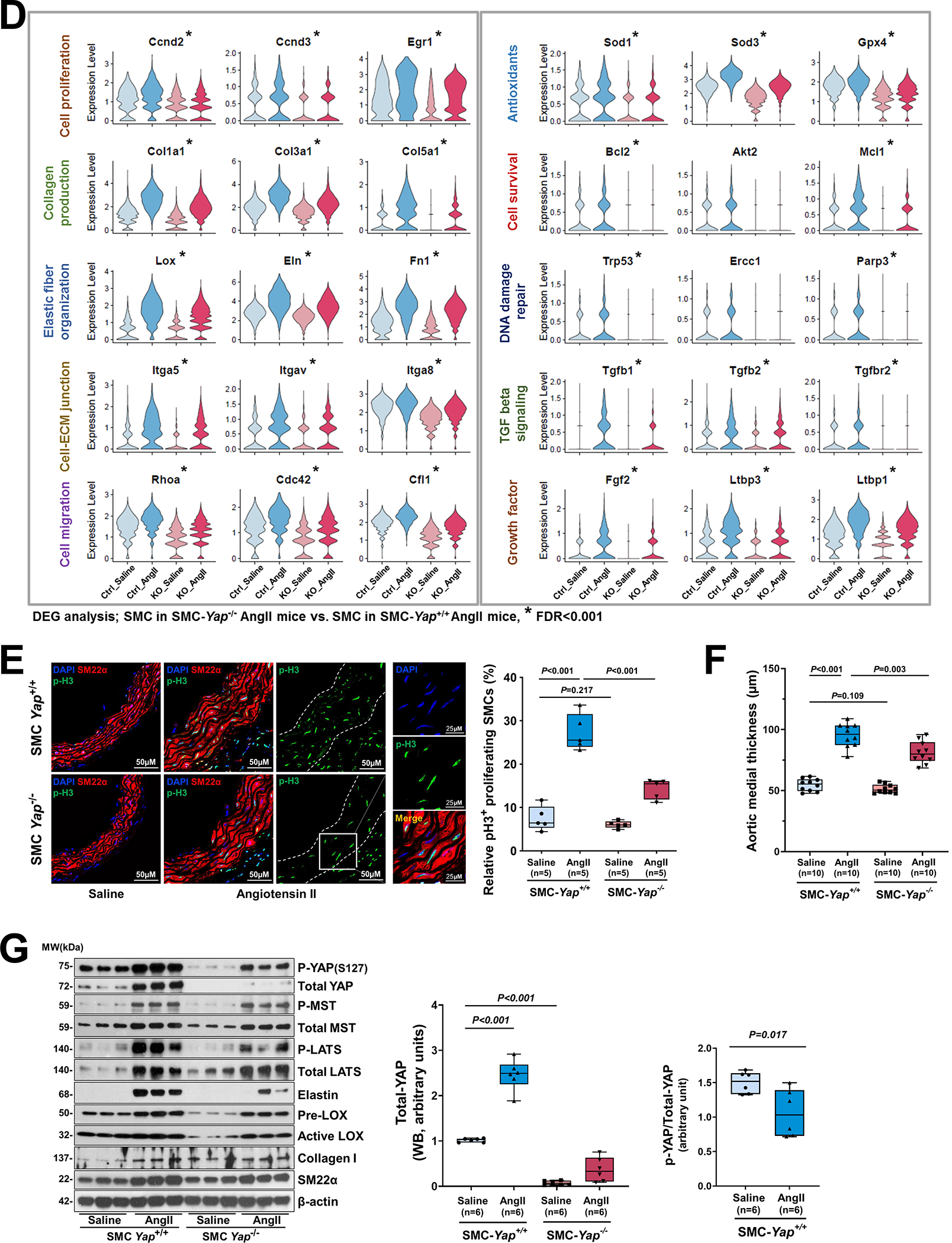
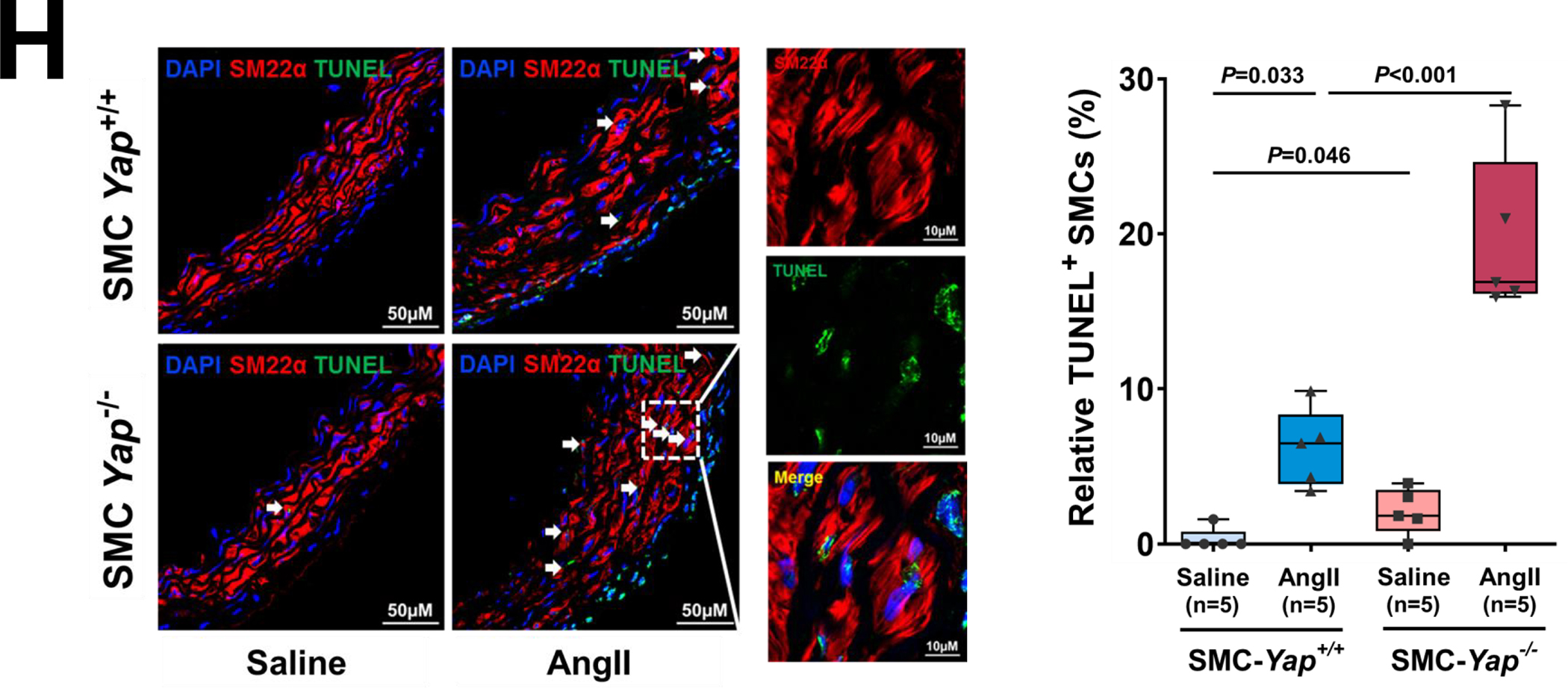
Results: In wild-type mice, AngII infusion increased medial thickness in the thoracic aorta. Single-cell RNA sequencing analysis revealed an adaptive response in thoracic SMCs characterized by upregulated genes with roles in wound healing, elastin and collagen production, proliferation, migration, cytoskeleton organization, cell-matrix focal adhesion, and PI3K-PKB/Akt (phosphoinositide-3-kinase-protein kinase B/Akt) and TGF-β (transforming growth factor beta) signaling. ScATAC-seq analysis showed increased chromatin accessibility at regulatory regions of adaptive genes and revealed the mechanical sensor YAP/transcriptional enhanced associate domains as a top candidate transcription complex driving the expression of these genes (eg, Lox, Col5a2, Tgfb2). In cultured human aortic SMCs, cyclic stretch activated YAP, which directly bound to adaptive gene regulatory regions (eg, Lox) and increased their transcript abundance. SMC-specific Yap deletion in mice compromised this adaptive response in SMCs, leading to an increased AAD incidence.
Conclusions: Aortic stress triggers the systemic epigenetic induction of an adaptive response (eg, wound healing, proliferation, matrix organization) in thoracic aortic SMCs that depends on functional biomechanical signal transduction (eg, YAP signaling). Our study highlights the importance of the adaptive response in maintaining aortic homeostasis and preventing AAD in mice.
Keywords: aortic aneurysm; extracellular matrix; homeostasis; mechanical stress; smooth muscle cell.
Conflict of interest statement
Disclosures
Dr. LeMaire serves as a consultant for Terumo Aortic and Cerus; and serves as a principal investigator for clinical studies sponsored by Terumo Aortic and CytoSorbents.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
