

SHP-2 and PD-1-SHP-2 signaling regulate myeloid cell differentiation and antitumor responses
- PMID: 36581713
- PMCID: PMC9810534
- DOI: 10.1038/s41590-022-01385-x
SHP-2 and PD-1-SHP-2 signaling regulate myeloid cell differentiation and antitumor responses
Abstract
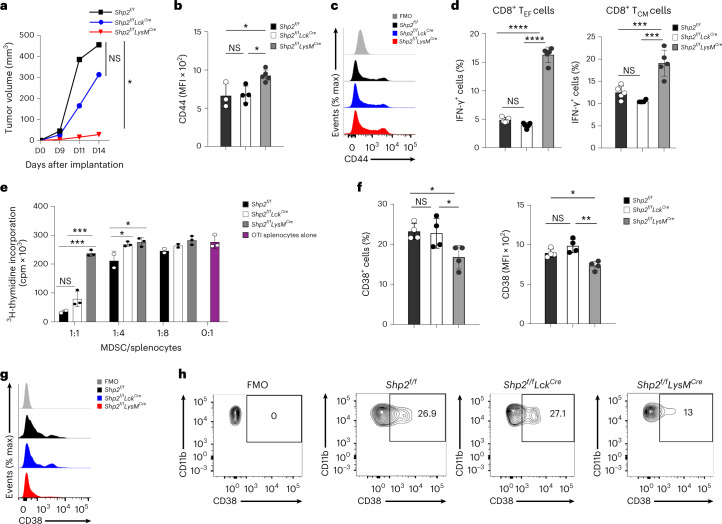
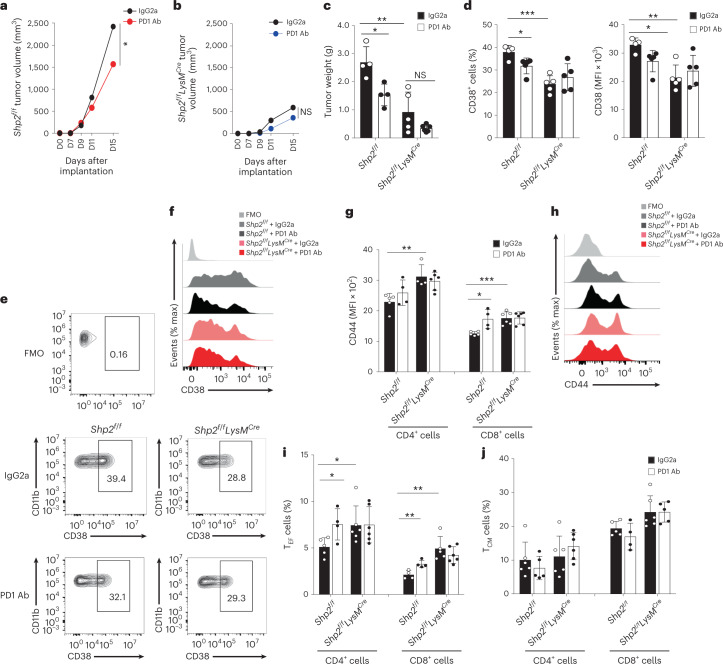
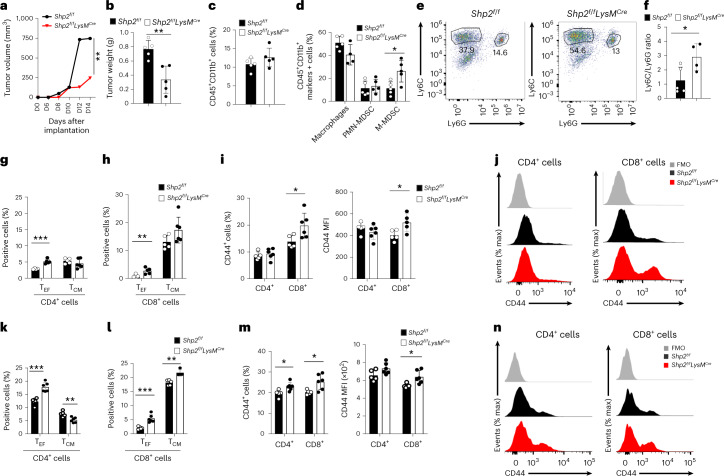
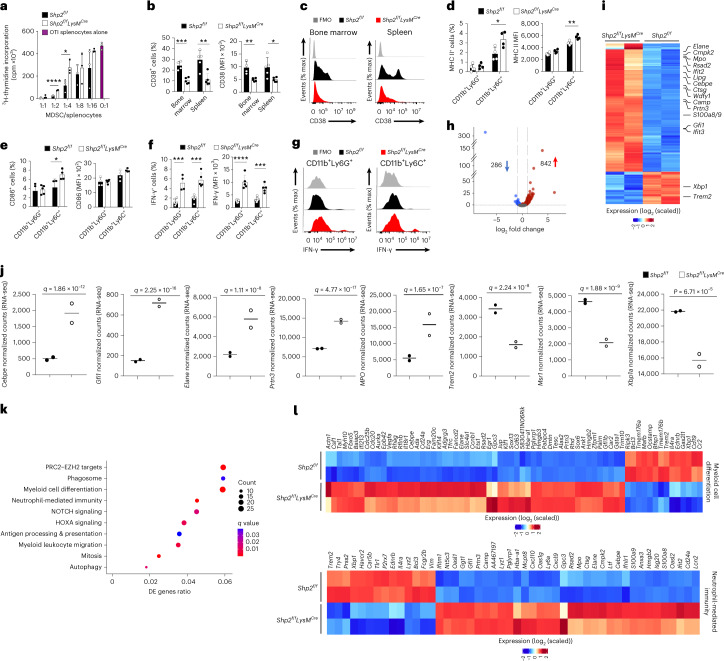
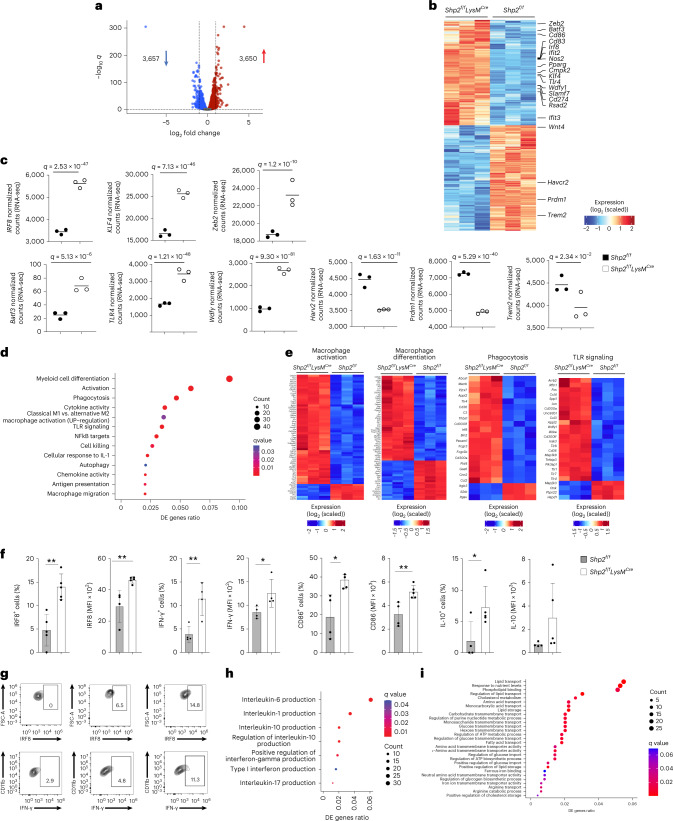
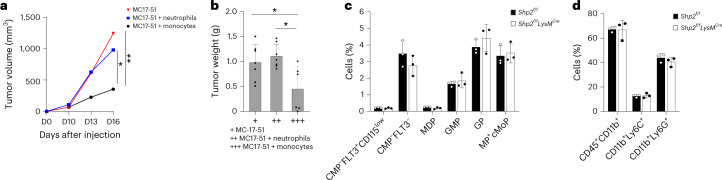
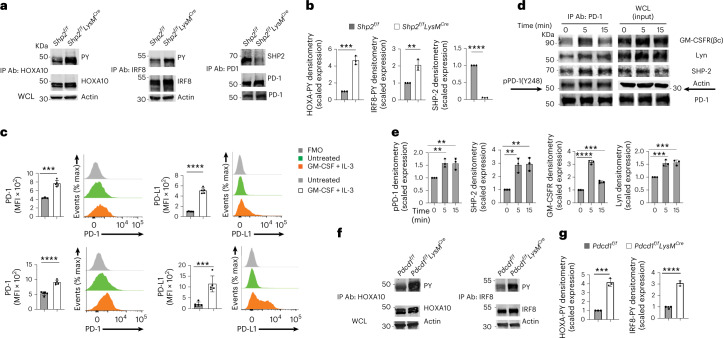
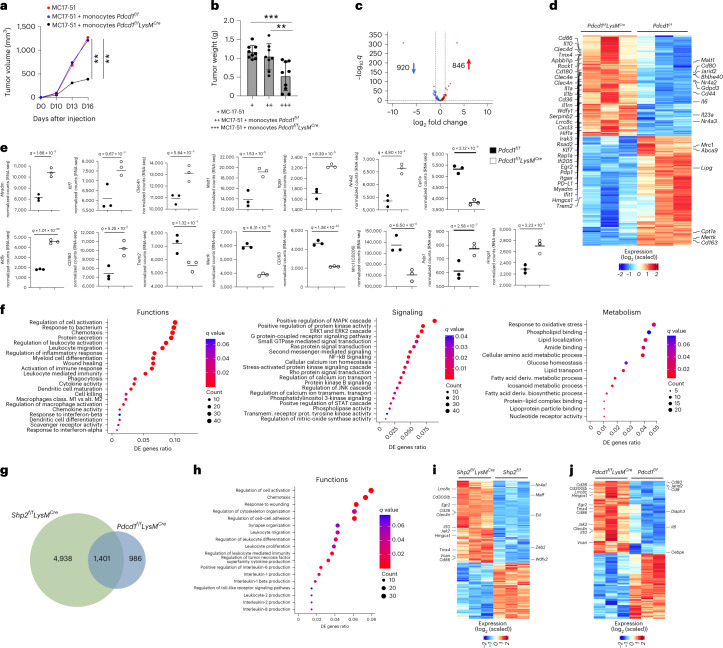
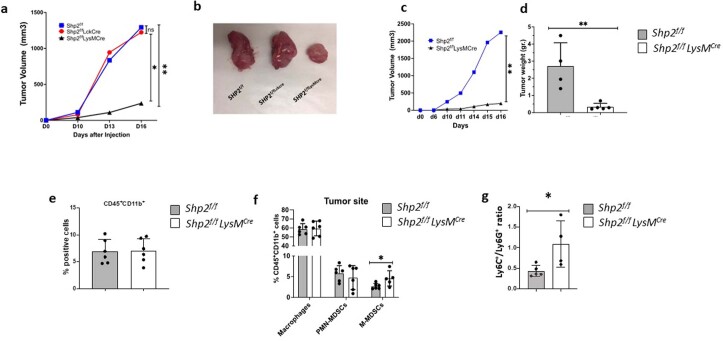
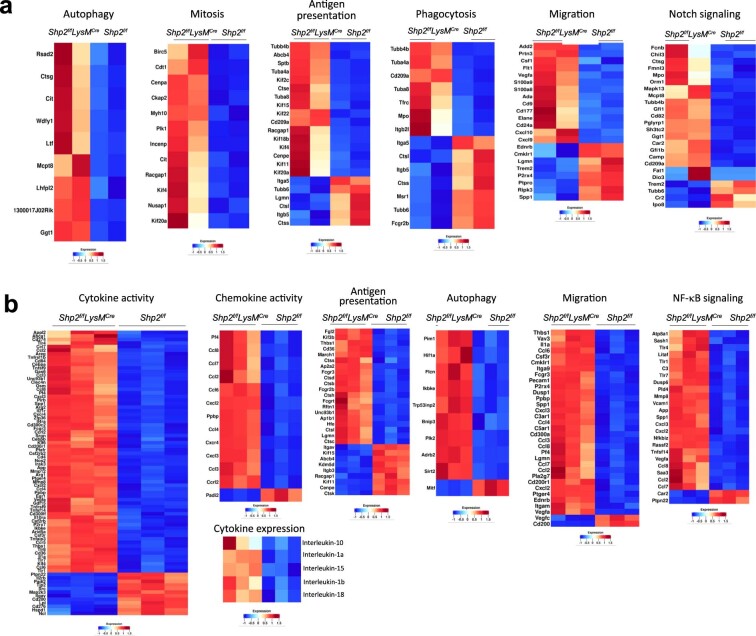
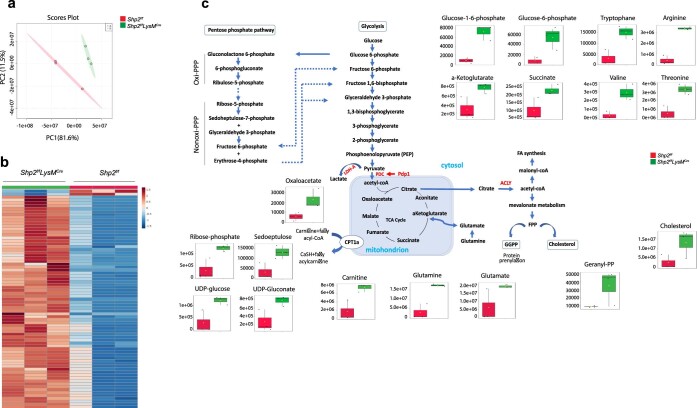
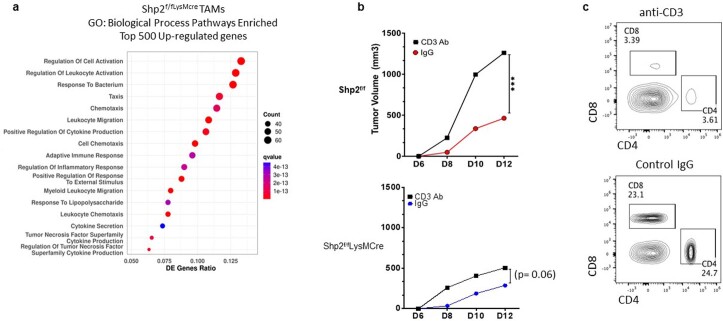
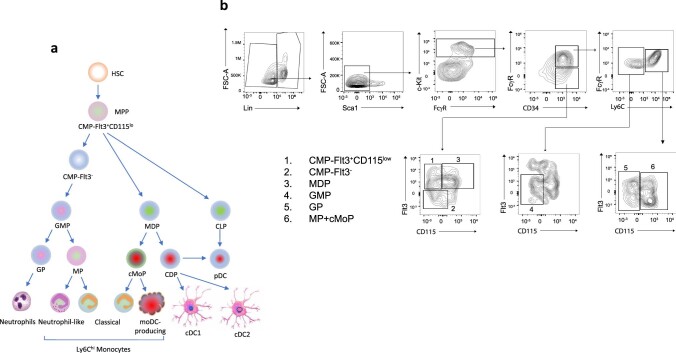
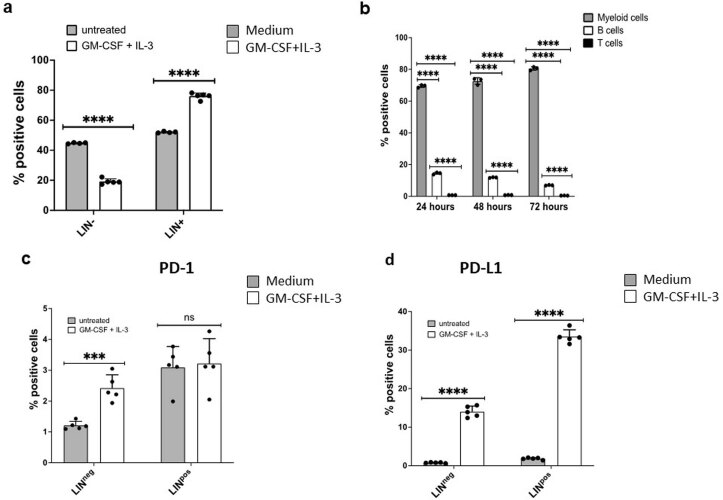
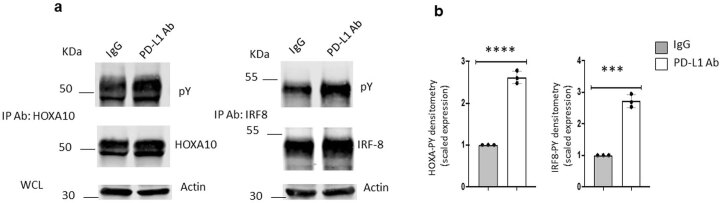
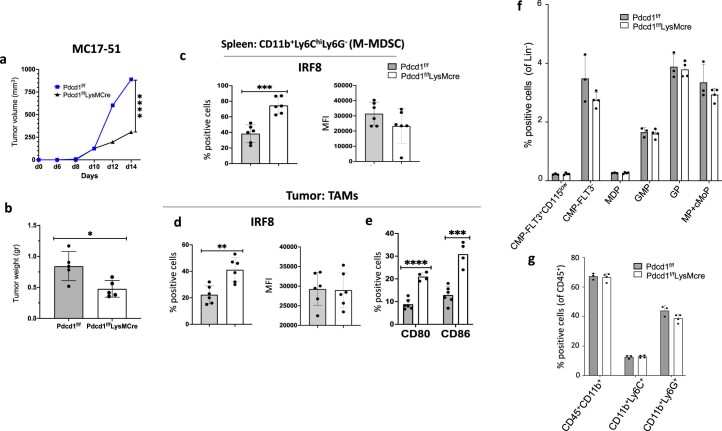
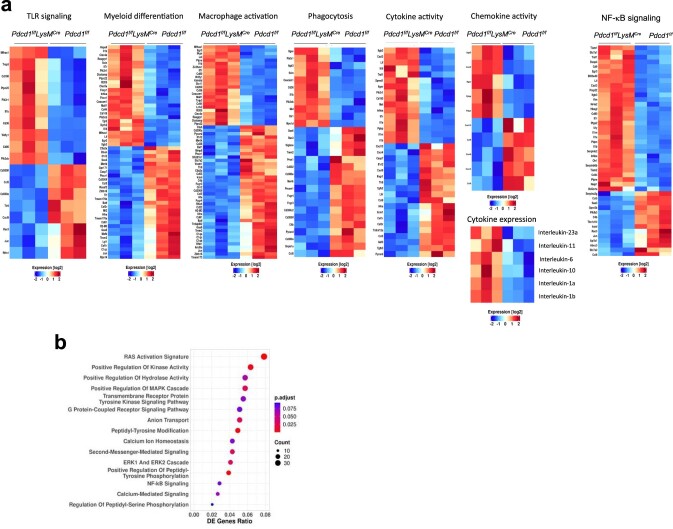
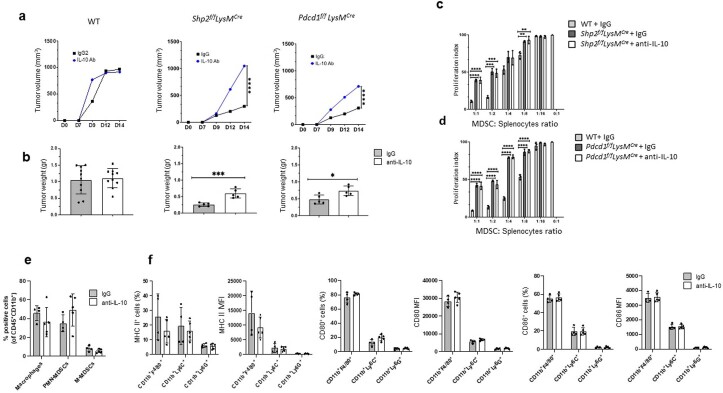
The inhibitory receptor PD-1 suppresses T cell activation by recruiting the phosphatase SHP-2. However, mice with a T-cell-specific deletion of SHP-2 do not have improved antitumor immunity. Here we showed that mice with conditional targeting of SHP-2 in myeloid cells, but not in T cells, had diminished tumor growth. RNA sequencing (RNA-seq) followed by gene set enrichment analysis indicated the presence of polymorphonuclear myeloid-derived suppressor cells and tumor-associated macrophages (TAMs) with enriched gene expression profiles of enhanced differentiation, activation and expression of immunostimulatory molecules. In mice with conditional targeting of PD-1 in myeloid cells, which also displayed diminished tumor growth, TAMs had gene expression profiles enriched for myeloid differentiation, activation and leukocyte-mediated immunity displaying >50% overlap with enriched profiles of SHP-2-deficient TAMs. In bone marrow, GM-CSF induced the phosphorylation of PD-1 and recruitment of PD-1-SHP-2 to the GM-CSF receptor. Deletion of SHP-2 or PD-1 enhanced GM-CSF-mediated phosphorylation of the transcription factors HOXA10 and IRF8, which regulate myeloid differentiation and monocytic-moDC lineage commitment, respectively. Thus, SHP-2 and PD-1-SHP-2 signaling restrained myelocyte differentiation resulting in a myeloid landscape that suppressed antitumor immunity.
© 2022. The Author(s).
Conflict of interest statement
V.A.B. has patents on the PD-1 pathway licensed by Bristol-Myers Squibb, Roche, Merck, EMD-Serono, Boehringer Ingelheim, AstraZeneca, Novartis and Dako. All the other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
