

Coordinated cadherin functions sculpt respiratory motor circuit connectivity
- PMID: 36583530
- PMCID: PMC9910829
- DOI: 10.7554/eLife.82116
Coordinated cadherin functions sculpt respiratory motor circuit connectivity
Abstract
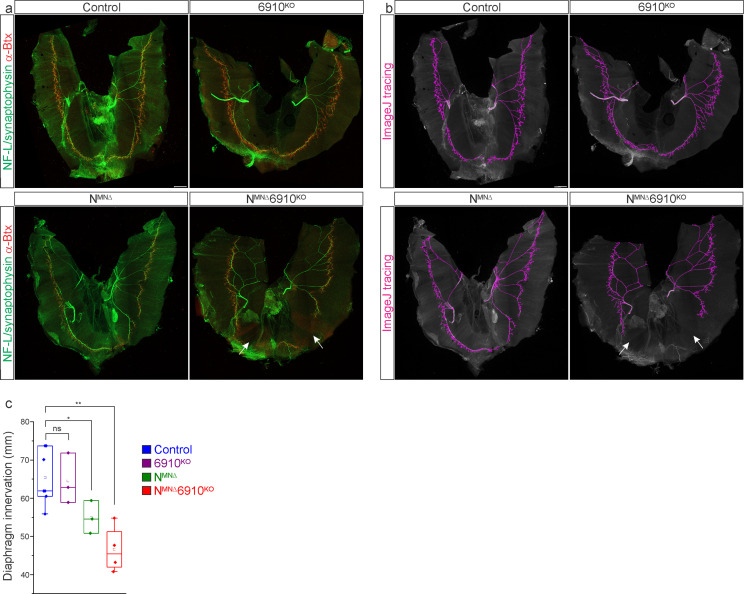
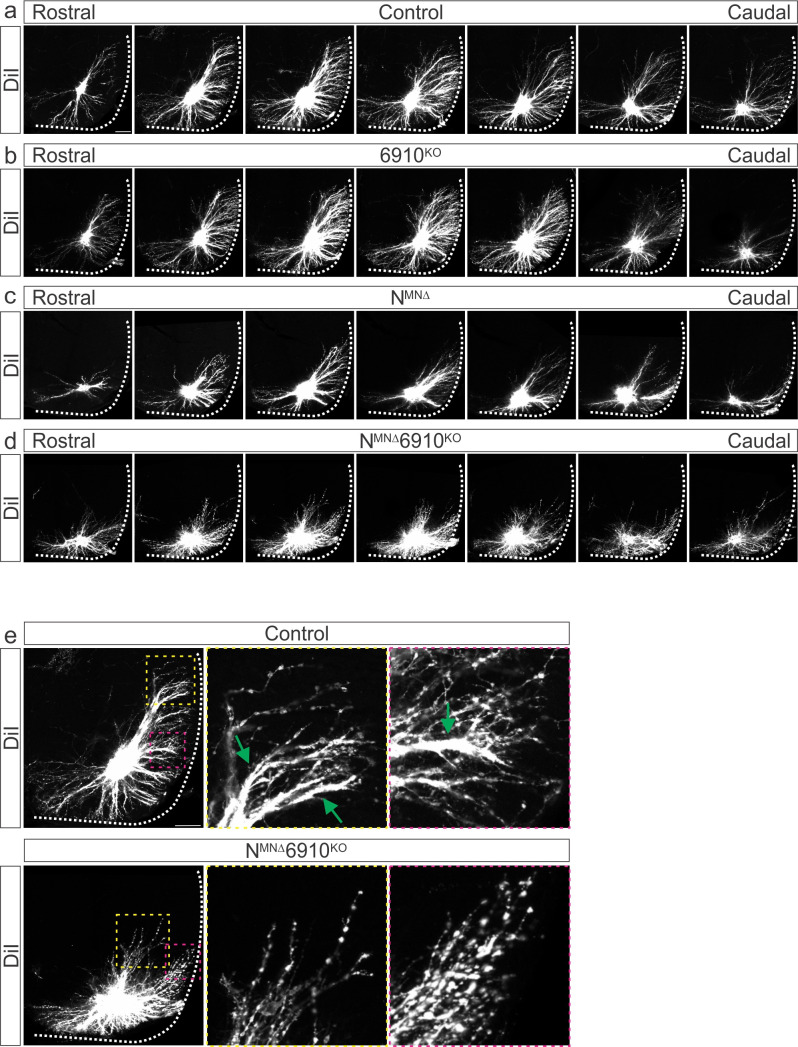
Breathing, and the motor circuits that control it, is essential for life. At the core of respiratory circuits are Dbx1-derived interneurons, which generate the rhythm and pattern of breathing, and phrenic motor neurons (MNs), which provide the final motor output that drives diaphragm muscle contractions during inspiration. Despite their critical function, the principles that dictate how respiratory circuits assemble are unknown. Here, we show that coordinated activity of a type I cadherin (N-cadherin) and type II cadherins (Cadherin-6, -9, and -10) is required in both MNs and Dbx1-derived neurons to generate robust respiratory motor output. Both MN- and Dbx1-specific cadherin inactivation in mice during a critical developmental window results in perinatal lethality due to respiratory failure and a striking reduction in phrenic MN bursting activity. This combinatorial cadherin code is required to establish phrenic MN cell body and dendritic topography; surprisingly, however, cell body position appears to be dispensable for the targeting of phrenic MNs by descending respiratory inputs. Our findings demonstrate that type I and II cadherins function cooperatively throughout the respiratory circuit to generate a robust breathing output and reveal novel strategies that drive the assembly of motor circuits.
Keywords: cadherins; developmental biology; mouse; neuroscience; phrenic motor neurons; rVRG; respiratory circuits.
Plain language summary
The neural circuits which control breathing are established in the womb, ready to switch on with the first gulp of air. Defects in the way that this network is assembled can result in conditions such as sudden infant death syndrome. This process, however, remains poorly understood; in particular, it is still unclear exactly how the two main types of nerve cells which form respiratory circuits start to ‘talk’ to each other. Known as Dbx1-derived interneurons and phrenic motor neurons, these cell populations reside in different parts of the body and perform distinct roles. The interneurons, which are present in the brainstem, act as a pacemaker to set the rhythm of respiration; the motor neurons reside in the spinal cord, connecting the interneurons with the muscles which allow the lungs to fill with air. Vagnozzi et al. aimed to identify how phrenic motor neurons connect to and relay signals from other neurons involved in breathing to the diaphragm muscle. To do so, the team focused on cadherins, a group of proteins which allow cells to attach to one another. Studded through the membrane, these molecules are also often involved in forming connections from one cell to another that allow them to communicate. Newborn mice in which phrenic motor neurons lacked a specific combination of cadherins experienced respiratory failure, showing that these proteins were needed for breathing circuits to develop normally. Electrical activity recorded from these cells showed that phrenic motor neurons lacking cadherins could not receive the signals required to activate the breathing muscles. Microscopy imaging also revealed that the loss of cadherins shifted the position of the phrenic motor neurons within the spinal cord; however, this change did not seem to affect the connections these cells could establish. The ability to breathe is compromised in many incurable human diseases such as muscular dystrophies and amyotrophic lateral sclerosis. It may be possible to alleviate some of these symptoms by integrating phrenic motor neurons created in the laboratory into existing circuits. Studies which aim to decipher how the respiratory network is established, such as the one conducted by Vagnozzi et al., are essential in this effort.
© 2022, Vagnozzi et al.
Conflict of interest statement
AV, MM, ML, EB, RK, AA, LS, XD, NZ, LL, PP No competing interests declared
Figures












References
-
- Basu R, Duan X, Taylor MR, Martin EA, Muralidhar S, Wang Y, Gangi-Wellman L, Das SC, Yamagata M, West PJ, Sanes JR, Williams ME. Heterophilic type II cadherins are required for high-magnitude synaptic potentiation in the hippocampus. Neuron. 2017;96:160–176. doi: 10.1016/j.neuron.2017.09.009. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
