

Metabolic adaptation supports enhanced macrophage efferocytosis in limited-oxygen environments
- PMID: 36584675
- PMCID: PMC9908853
- DOI: 10.1016/j.cmet.2022.12.005
Metabolic adaptation supports enhanced macrophage efferocytosis in limited-oxygen environments
Abstract
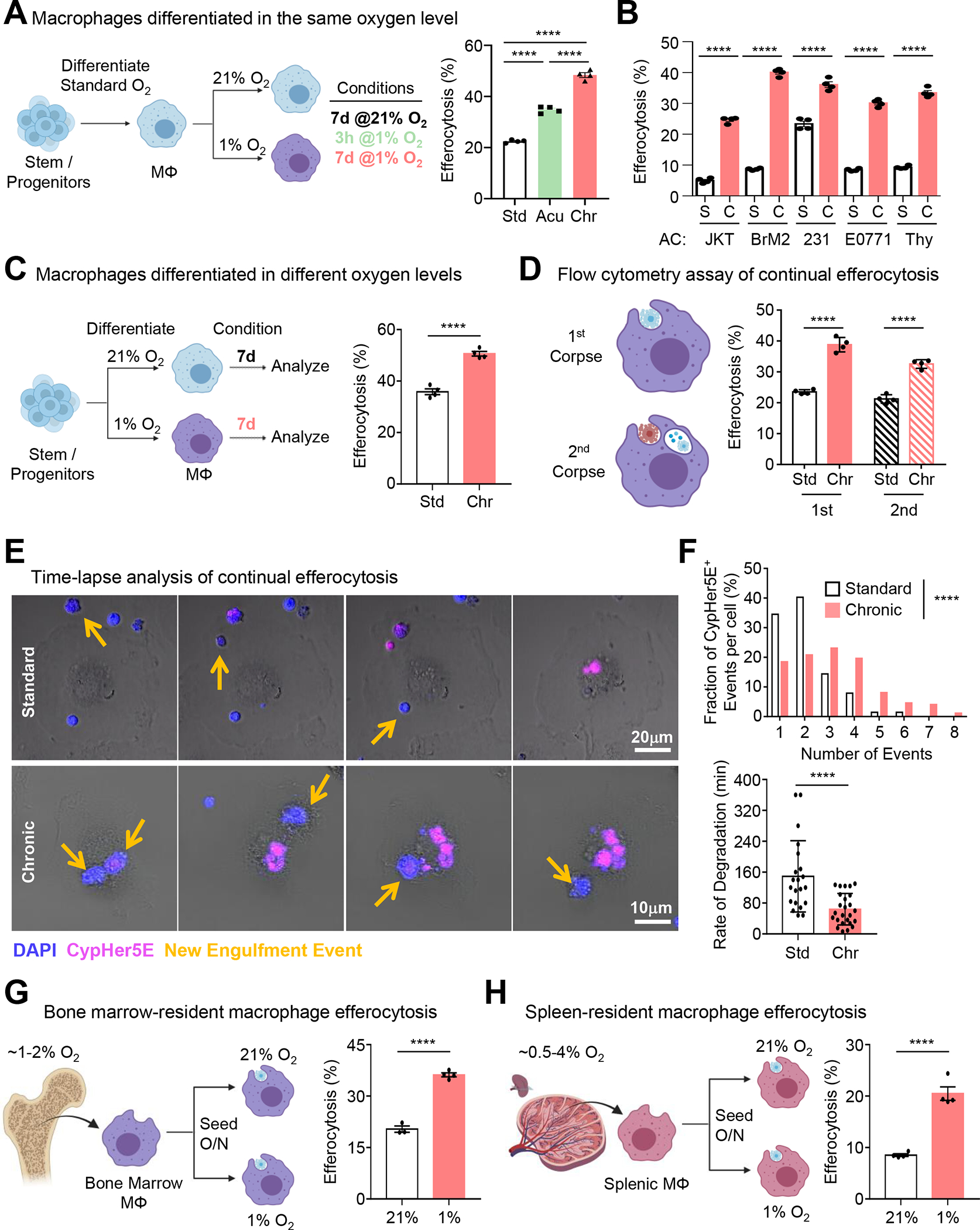
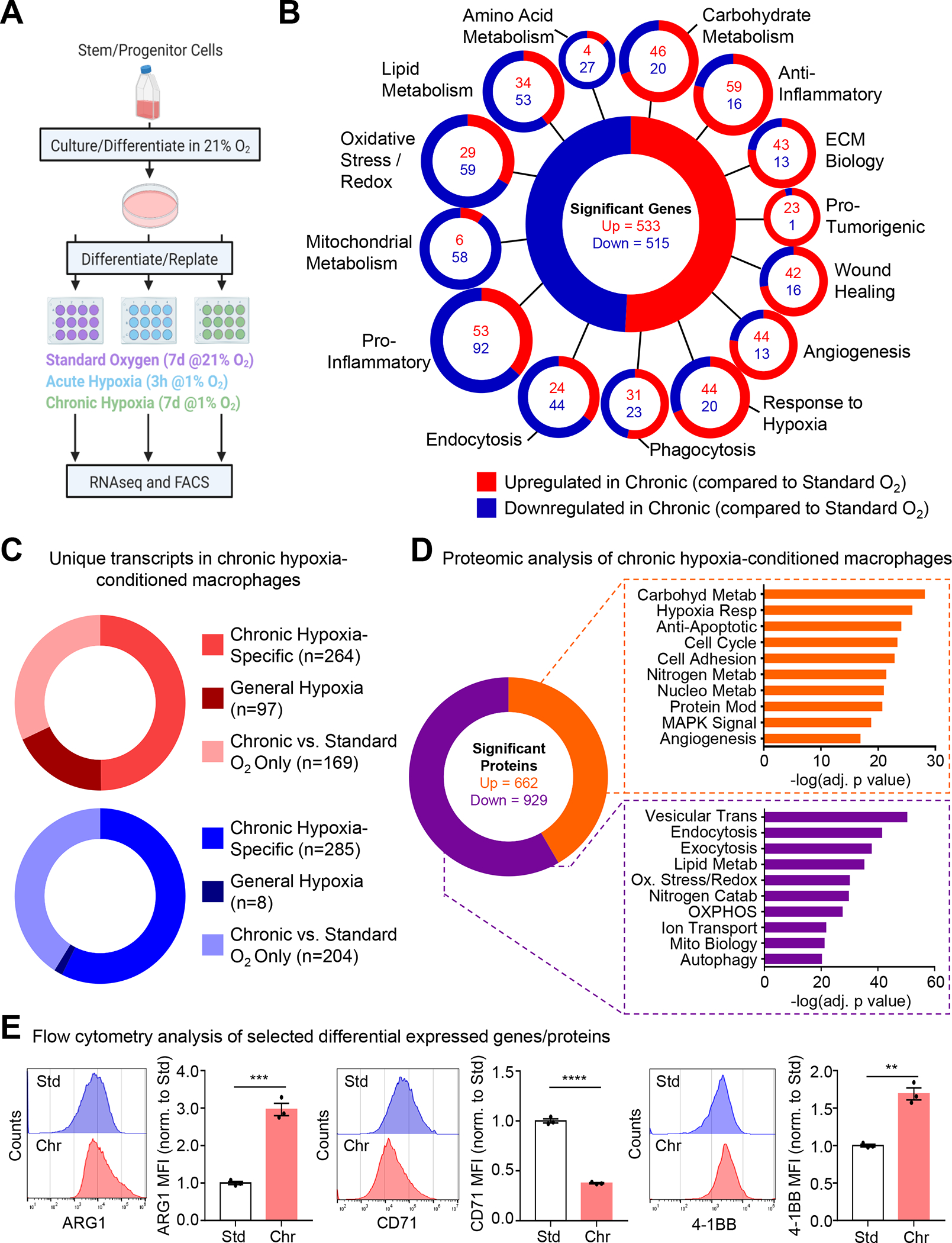
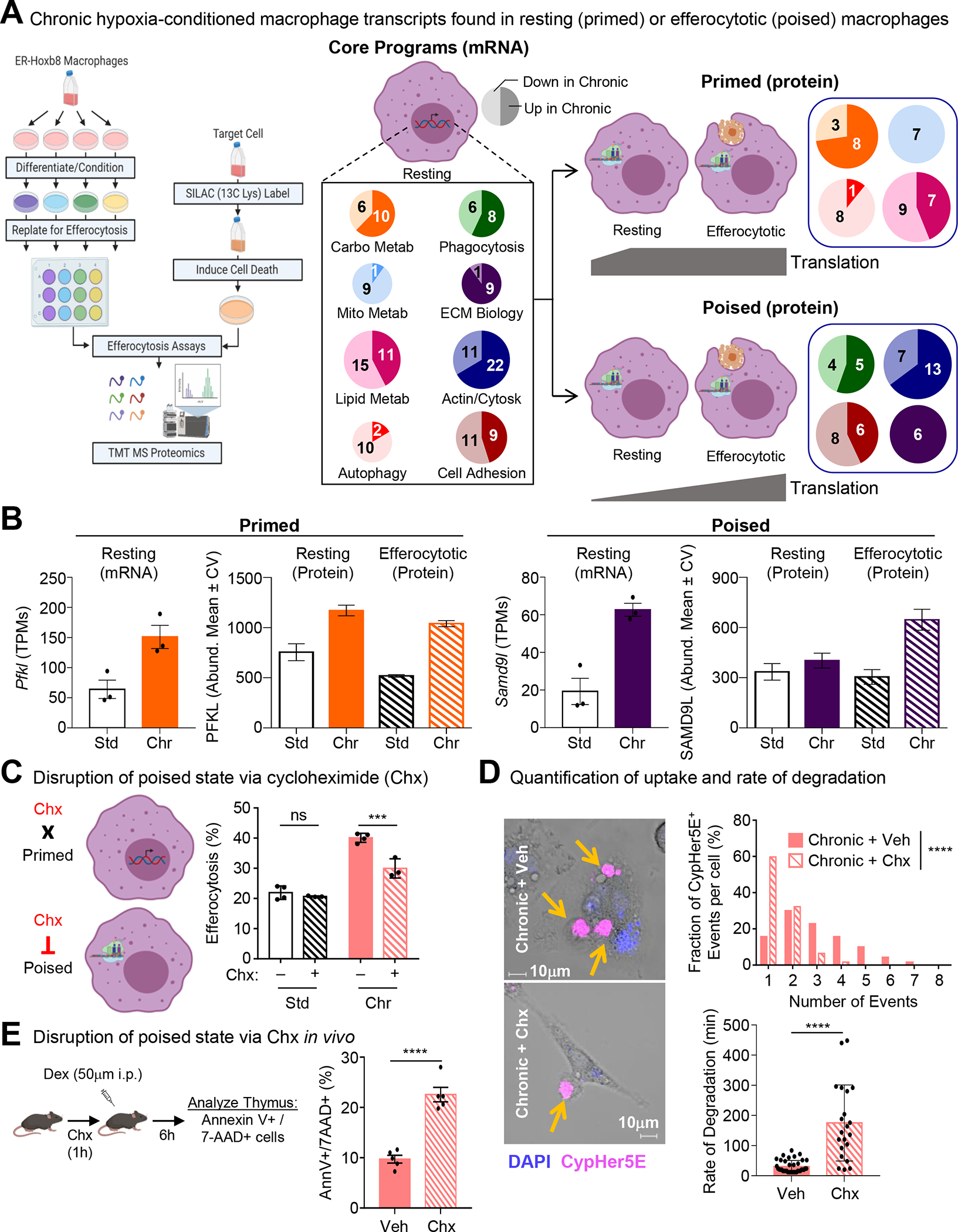
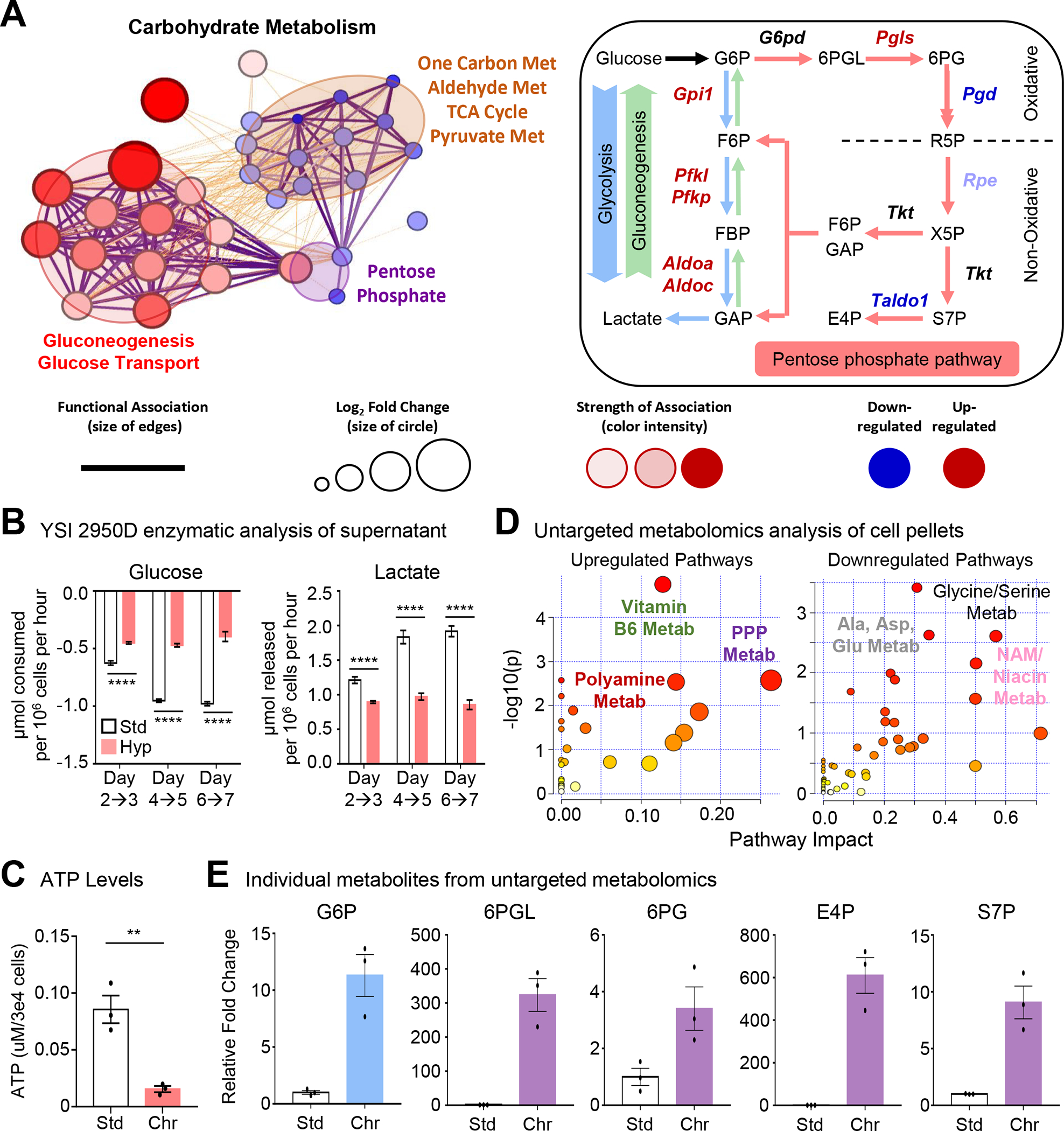
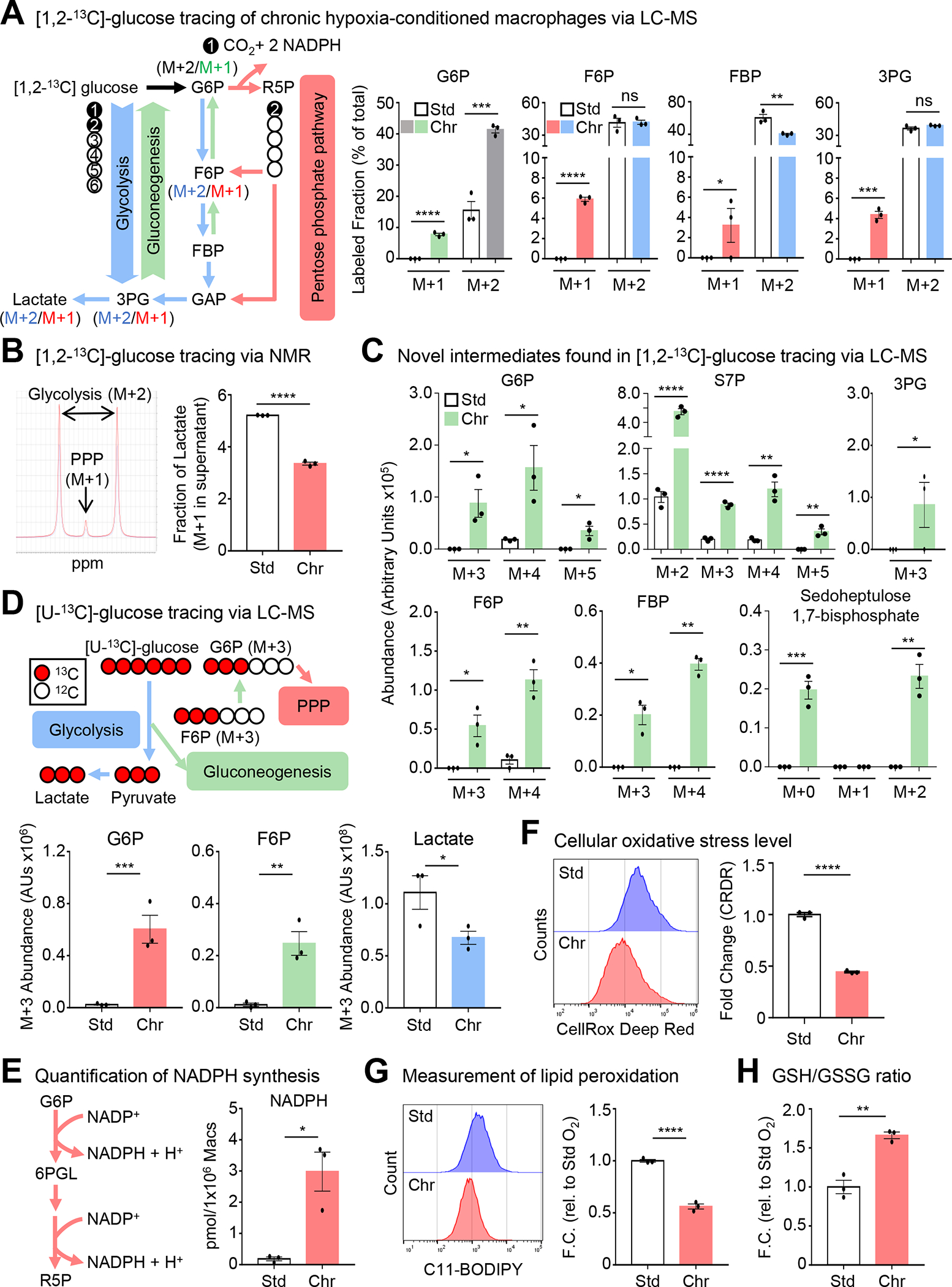
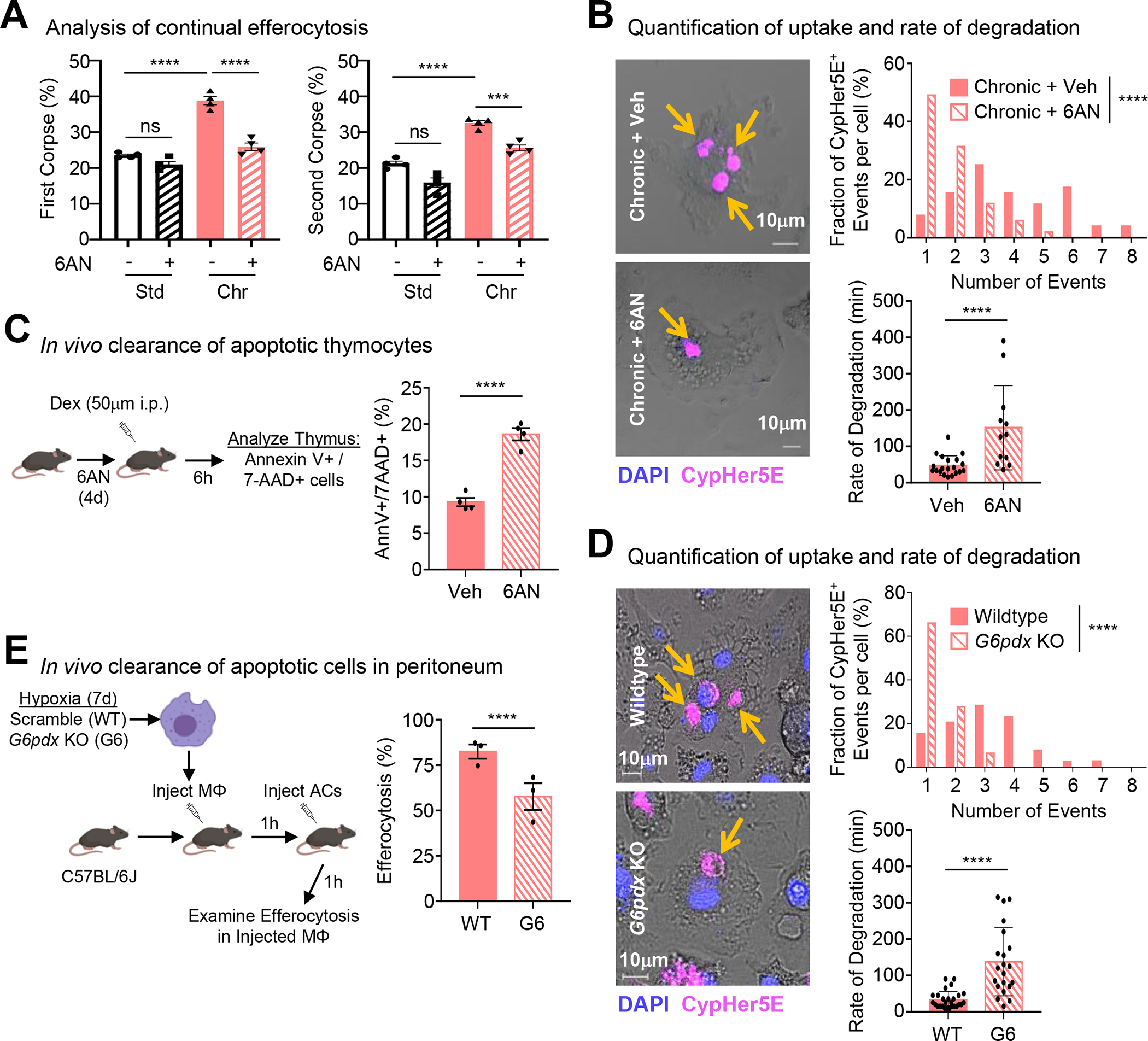
Apoptotic cell (AC) clearance (efferocytosis) is performed by phagocytes, such as macrophages, that inhabit harsh physiological environments. Here, we find that macrophages display enhanced efferocytosis under prolonged (chronic) physiological hypoxia, characterized by increased internalization and accelerated degradation of ACs. Transcriptional and translational analyses revealed that chronic physiological hypoxia induces two distinct but complimentary states. The first, "primed" state, consists of concomitant transcription and translation of metabolic programs in AC-naive macrophages that persist during efferocytosis. The second, "poised" state, consists of transcription, but not translation, of phagocyte function programs in AC-naive macrophages that are translated during efferocytosis. Mechanistically, macrophages efficiently flux glucose into a noncanonical pentose phosphate pathway (PPP) loop to enhance NADPH production. PPP-derived NADPH directly supports enhanced efferocytosis under physiological hypoxia by ensuring phagolysosomal maturation and redox homeostasis. Thus, macrophages residing under physiological hypoxia adopt states that support cell fitness and ensure performance of essential homeostatic functions rapidly and safely.
Keywords: apoptotic cell clearance; cellular adaptation; efferocytosis; homeostasis; metabolism; oxygen; pentose phosphate pathway; physiological hypoxia.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests K.R.K. serves on the scientific advisory board of NVision Imaging Technologies. J.S.A.P. and K.R.K. holds patents related to imaging and modulation of cellular metabolism.
Figures

Similar articles
-
Pentose Phosphate Pathway Regulates Tolerogenic Apoptotic Cell Clearance and Immune Tolerance.Front Immunol. 2022 Jan 10;12:797091. doi: 10.3389/fimmu.2021.797091. eCollection 2021. Front Immunol. 2022. PMID: 35082786 Free PMC article.
-
Multiomics reveal the central role of pentose phosphate pathway in resident thymic macrophages to cope with efferocytosis-associated stress.Cell Rep. 2022 Jul 12;40(2):111065. doi: 10.1016/j.celrep.2022.111065. Cell Rep. 2022. PMID: 35830797
-
NADPH oxidase activation regulates apoptotic neutrophil clearance by murine macrophages.Blood. 2018 May 24;131(21):2367-2378. doi: 10.1182/blood-2017-09-809004. Epub 2018 Apr 4. Blood. 2018. PMID: 29618478 Free PMC article.
-
Mechanism of Efferocytosis in Determining Ischaemic Stroke Resolution-Diving into Microglia/Macrophage Functions and Therapeutic Modality.Mol Neurobiol. 2024 Oct;61(10):7583-7602. doi: 10.1007/s12035-024-04060-4. Epub 2024 Feb 27. Mol Neurobiol. 2024. PMID: 38409642 Review.
-
Efferocytosis by macrophages in physiological and pathological conditions: regulatory pathways and molecular mechanisms.Front Immunol. 2024 May 8;15:1275203. doi: 10.3389/fimmu.2024.1275203. eCollection 2024. Front Immunol. 2024. PMID: 38779685 Free PMC article. Review.
Cited by
-
circROBO1 promotes prostate cancer growth and enzalutamide resistance via accelerating glycolysis.J Cancer. 2023 Aug 21;14(13):2574-2584. doi: 10.7150/jca.86940. eCollection 2023. J Cancer. 2023. PMID: 37670963 Free PMC article.
-
Metabolic engineering to facilitate anti-tumor immunity.Cancer Cell. 2025 Mar 10;43(3):552-562.e9. doi: 10.1016/j.ccell.2025.02.004. Epub 2025 Feb 27. Cancer Cell. 2025. PMID: 40020672
-
FOLR2+ macrophages in cancer: allies or enemies.Cell Commun Signal. 2025 Jun 2;23(1):261. doi: 10.1186/s12964-025-02257-1. Cell Commun Signal. 2025. PMID: 40457348 Free PMC article. Review.
-
Disulfidptosis: A new type of cell death.Apoptosis. 2024 Oct;29(9-10):1309-1329. doi: 10.1007/s10495-024-01989-8. Epub 2024 Jun 17. Apoptosis. 2024. PMID: 38886311 Free PMC article. Review.
-
Molecular dynamics of inflammation resolution: therapeutic implications.Front Cell Dev Biol. 2025 May 8;13:1600149. doi: 10.3389/fcell.2025.1600149. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40406415 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
