

A delicate balance between antibody evasion and ACE2 affinity for Omicron BA.2.75
- PMID: 36586406
- PMCID: PMC9747698
- DOI: 10.1016/j.celrep.2022.111903
A delicate balance between antibody evasion and ACE2 affinity for Omicron BA.2.75
Abstract
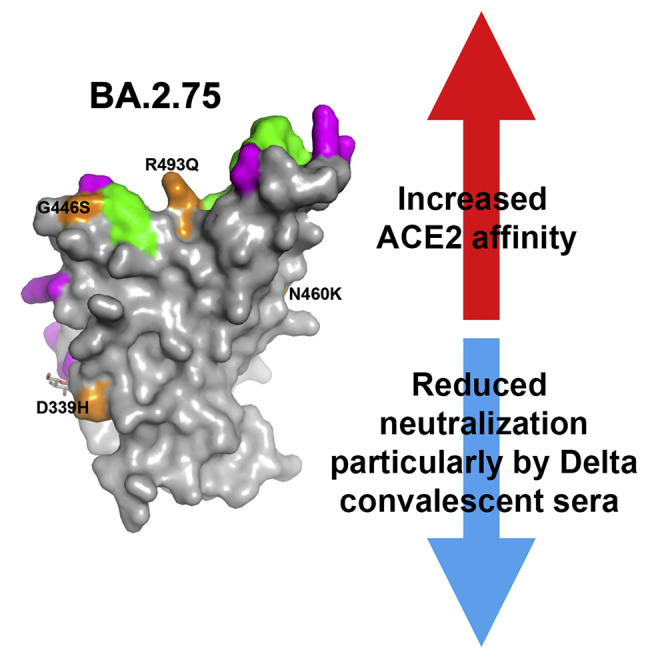
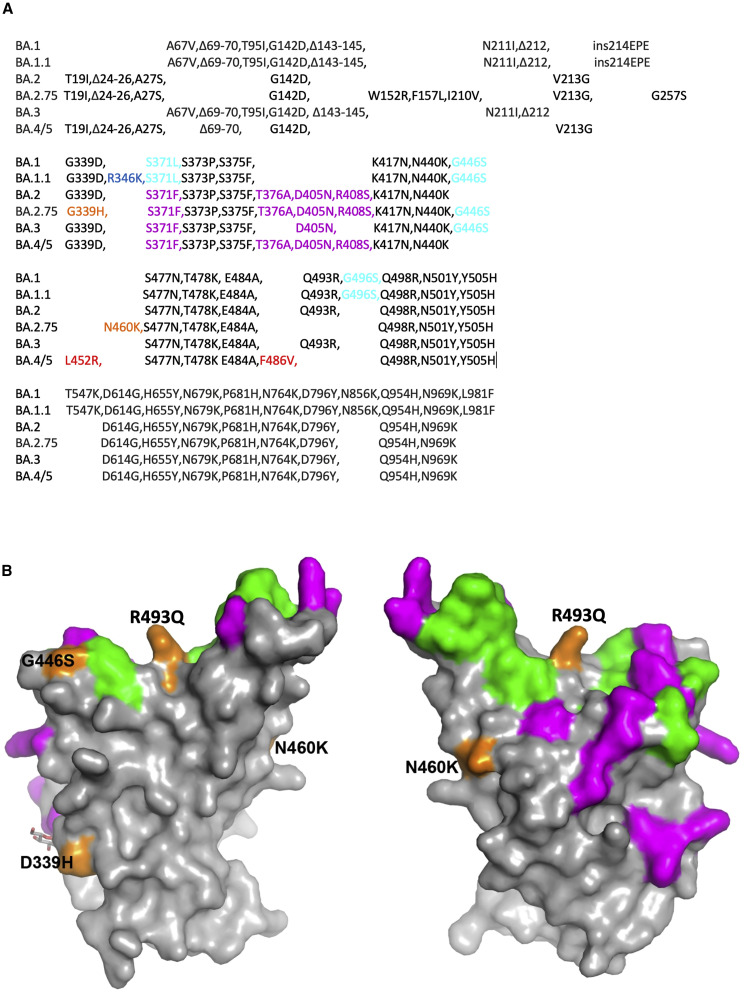
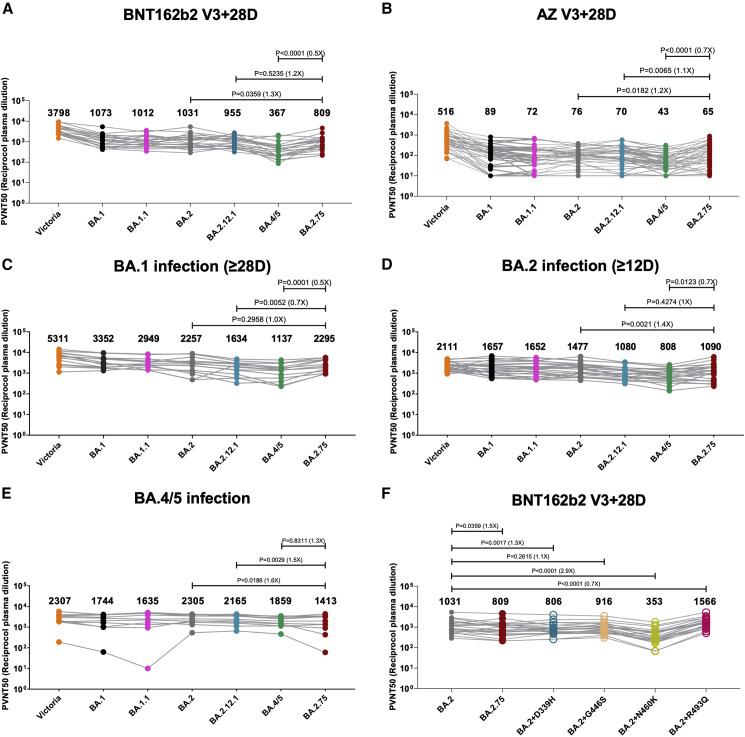
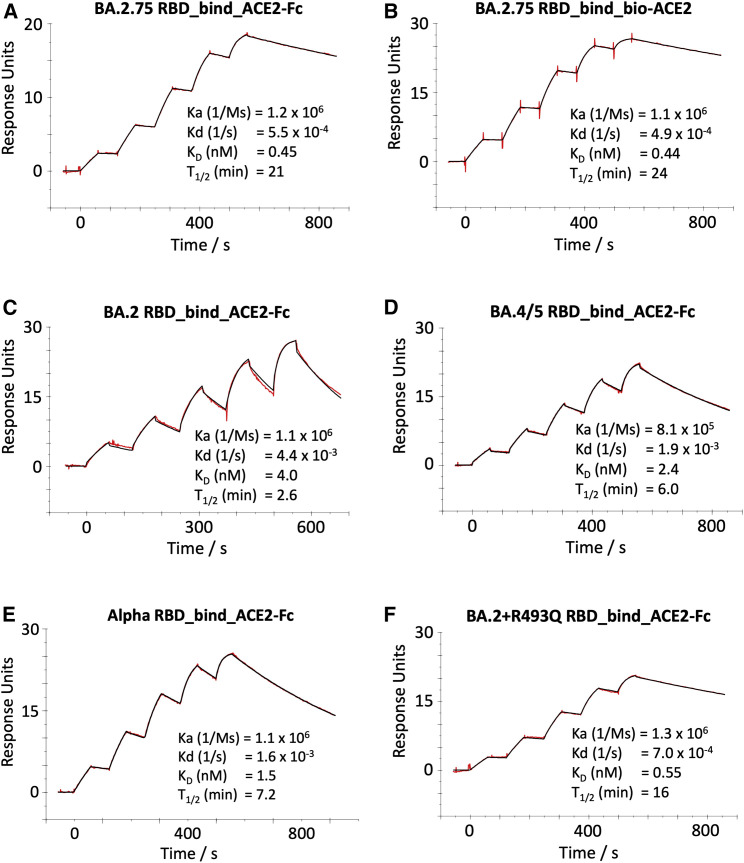
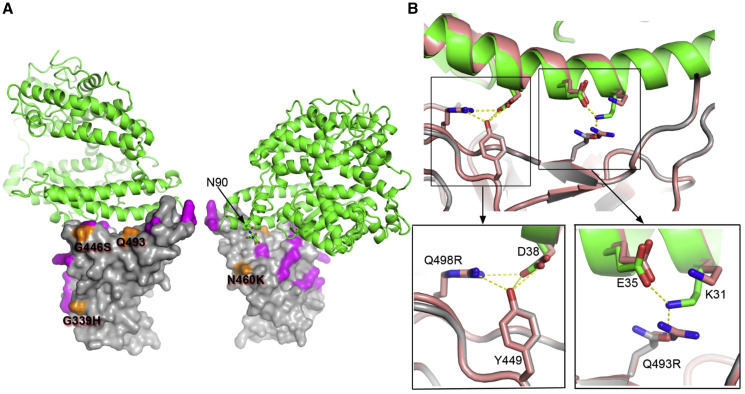
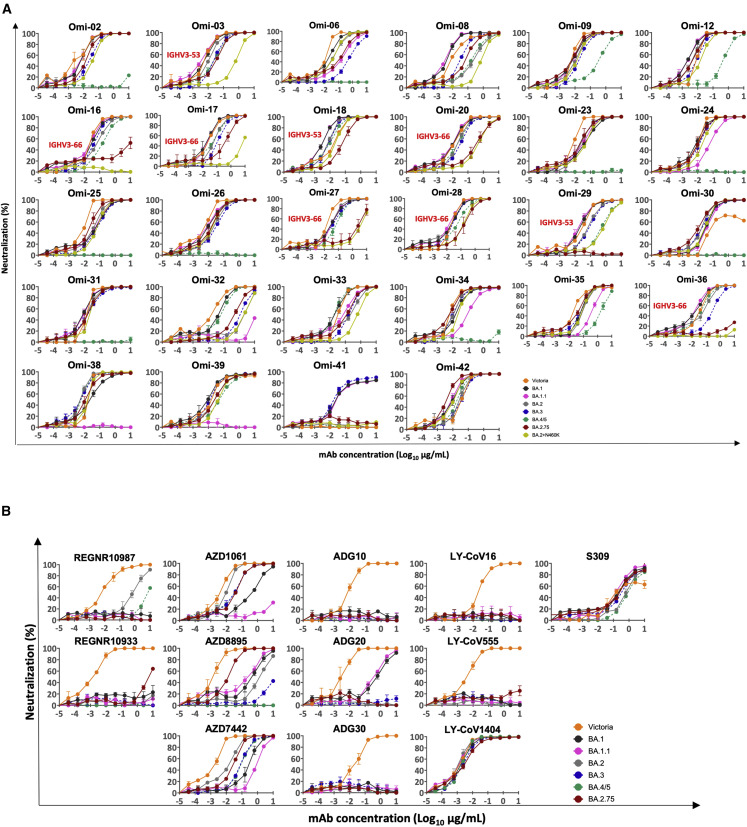
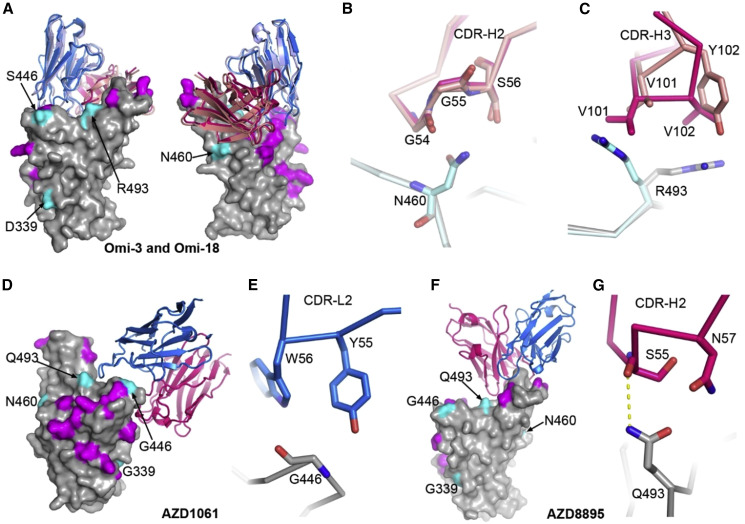
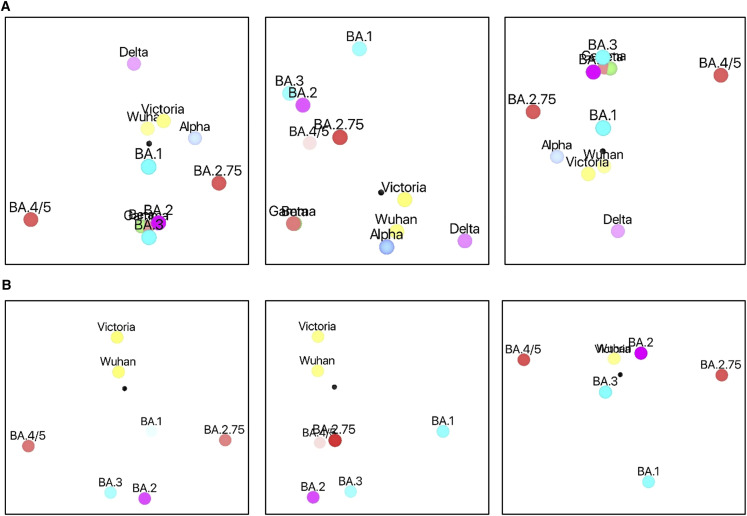
Variants of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) have caused successive global waves of infection. These variants, with multiple mutations in the spike protein, are thought to facilitate escape from natural and vaccine-induced immunity and often increase in affinity for ACE2. The latest variant to cause concern is BA.2.75, identified in India where it is now the dominant strain, with evidence of wider dissemination. BA.2.75 is derived from BA.2 and contains four additional mutations in the receptor-binding domain (RBD). Here, we perform an antigenic and biophysical characterization of BA.2.75, revealing an interesting balance between humoral evasion and ACE2 receptor affinity. ACE2 affinity for BA.2.75 is increased 9-fold compared with BA.2; there is also evidence of escape of BA.2.75 from immune serum, particularly that induced by Delta infection, which may explain the rapid spread in India, where where there is a high background of Delta infection. ACE2 affinity appears to be prioritized over greater escape.
Keywords: ACE2 receptor; BA.2.75; COVID-19; CP: Immunology; CP: Microbiology; RBD; SARS-CoV-2; antigenic variation; immune escape; spike; variant; variant of concern.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.R.S. sits on the GSK Vaccines Scientific Advisory Board, consults for Astra Zeneca, and is a founding member of RQ Biotechnology. Oxford University holds intellectual property related to the Oxford-Astra Zeneca vaccine and SARS-CoV-2 mAbs discovered in G.R.S.’s laboratory. A.J.P. is Chair of UK Dept. Health and Social Care’s (DHSC) Joint Committee on Vaccination & Immunisation (JCVI) but does not participate in the JCVI COVID-19 committee and is a member of the WHO’s SAGE. The views expressed in this article do not necessarily represent the views of DHSC, JCVI, or WHO. The University of Oxford has entered into a partnership with AstraZeneca on coronavirus vaccine development. T.L. is named as an inventor on a patent application covering this SARS-CoV-2 vaccine and was a consultant to Vaccitech for an unrelated project whilst the study was conducted. S.J.D. is a scientific advisor to the Scottish Parliament on COVID-19.
Figures
References
-
- Greninger A.L., Dien Bard J., Colgrove R.C., Graf E.H., Hanson K.E., Hayden M.K., Humphries R.M., Lowe C.F., Miller M.B., Pillai D.R., et al. Clinical and infection prevention applications of severe acute respiratory syndrome coronavirus 2 genotyping: an infectious diseases society of America/American society for microbiology consensus review document. J. Clin. Microbiol. 2022;60:e0165921. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
