

Infertile human endometrial organoid apical protein secretions are dysregulated and impair trophoblast progenitor cell adhesion
- PMID: 36589798
- PMCID: PMC9794621
- DOI: 10.3389/fendo.2022.1067648
Infertile human endometrial organoid apical protein secretions are dysregulated and impair trophoblast progenitor cell adhesion
Abstract
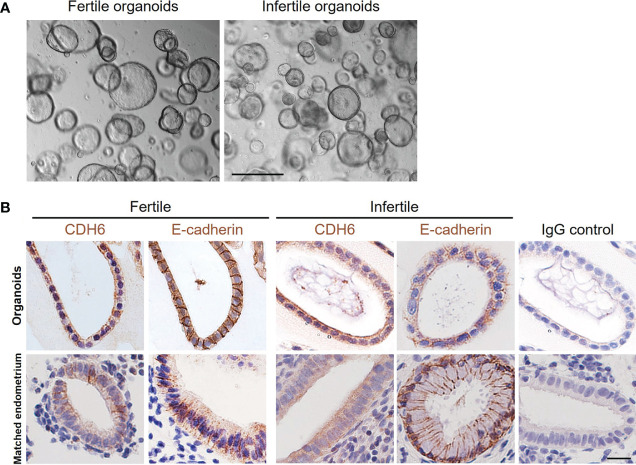
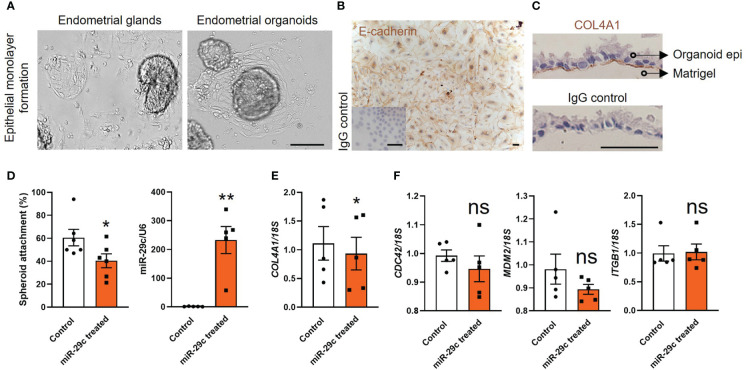
Introduction: Embryo implantation failure leads to infertility. As an important approach to regulate implantation, endometrial epithelial cells produce and secrete factors apically into the uterine cavity in the receptive phase to prepare the initial blastocyst adhesion and implantation. Organoids were recently developed from human endometrial epithelium with similar apical-basal polarity compared to endometrial gland making it an ideal model to study endometrial epithelial secretions.
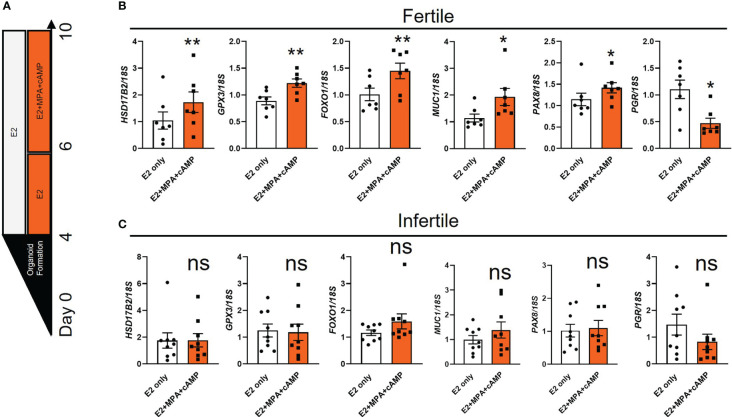
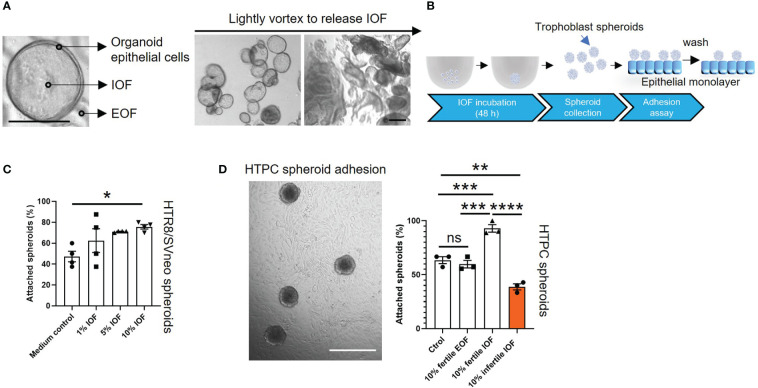
Methods: Endometrial organoids were established using endometrial biopsies from women with primary infertility and normal fertility. Fertile and infertile organoids were treated with hormones to model receptive phase of the endometrial epithelium and intra-organoid fluid (IOF) was collected to compare the apical protein secretion profile and function on trophoblast cell adhesion.
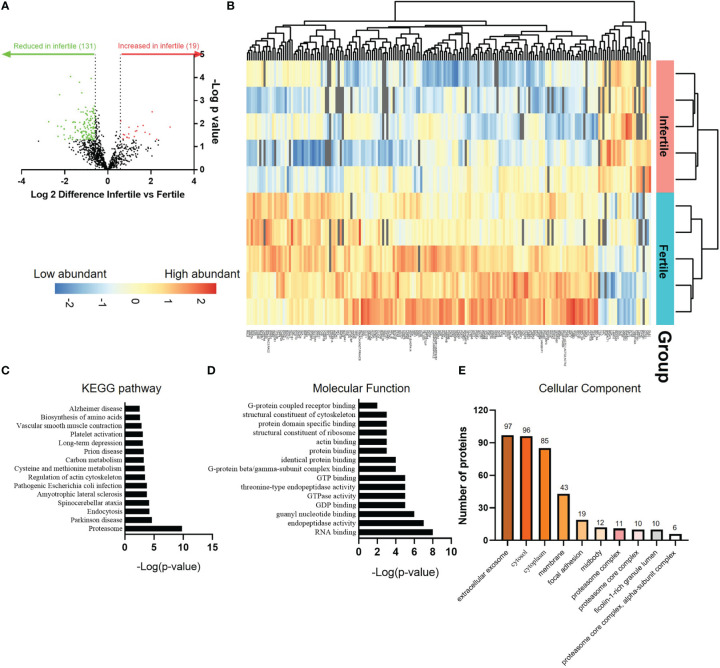
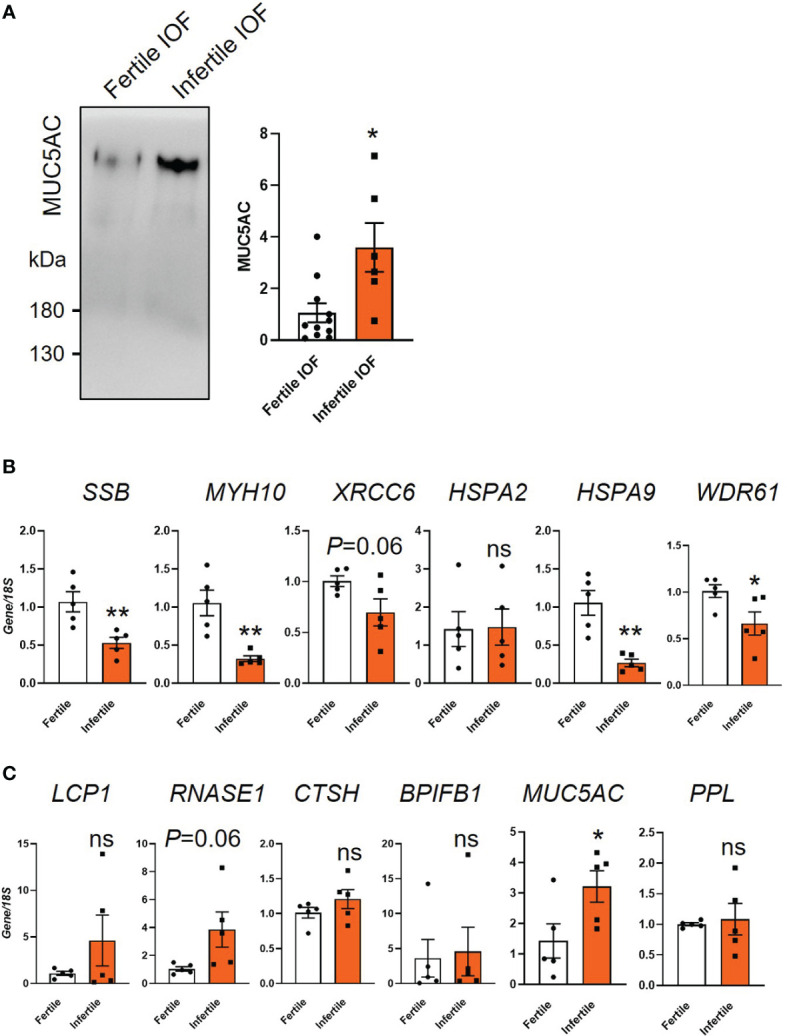
Results: Our data show that infertile organoids were dysregulated in their response to estrogen and progesterone treatment. Proteomic analysis of organoid apical secretions identified 150 dysregulated proteins between fertile and infertile groups (>1.5-fold change). Trophoblast progenitor spheroids (blastocyst surrogates) treated with infertile organoid apical secretions significantly compromised their adhesion to organoid epithelial cell monolayers compared to fertile group (P < 0.0001).
Discussion: This study revealed that endometrial organoid apical secretions alter trophoblast cell adhesiveness relative to fertility status of women. It paves the way to determine the molecular mechanisms by which endometrial epithelial apical released factors regulate blastocyst initial attachment and implantation.
Keywords: apical secretion; blastocyst attachment; endometrial organoids; infertile organoids; intra-organoid fluid.
Copyright © 2022 Zhou, Barton, Cui, Santos, Yang, Stern, Kieu, Teh, Ang, Lucky, Sgroi, Ye and Dimitriadis.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Salamun V, Bokal EV, Maver A, Papler TB. Transcriptome study of receptive endometrium in overweight and obese women shows important expression differences in immune response and inflammatory pathways in women who do not conceive. PLos One (2021) 16:e0261873. doi: 10.1371/journal.pone.0261873 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials