

Metabolic reprogramming of hepatocytes by Schistosoma mansoni eggs
- PMID: 36590323
- PMCID: PMC9800334
- DOI: 10.1016/j.jhepr.2022.100625
Metabolic reprogramming of hepatocytes by Schistosoma mansoni eggs
Abstract
Background & aims: Schistosomiasis is a parasitic infection which affects more than 200 million people globally. Schistosome eggs, but not the adult worms, are mainly responsible for schistosomiasis-specific morbidity in the liver. It is unclear if S. mansoni eggs consume host metabolites, and how this compromises the host parenchyma.
Methods: Metabolic reprogramming was analyzed by matrix-assisted laser desorption/ionization mass spectrometry imaging, liquid chromatography with high-resolution mass spectrometry, metabolite quantification, confocal laser scanning microscopy, live cell imaging, quantitative real-time PCR, western blotting, assessment of DNA damage, and immunohistology in hamster models and functional experiments in human cell lines. Major results were validated in human biopsies.
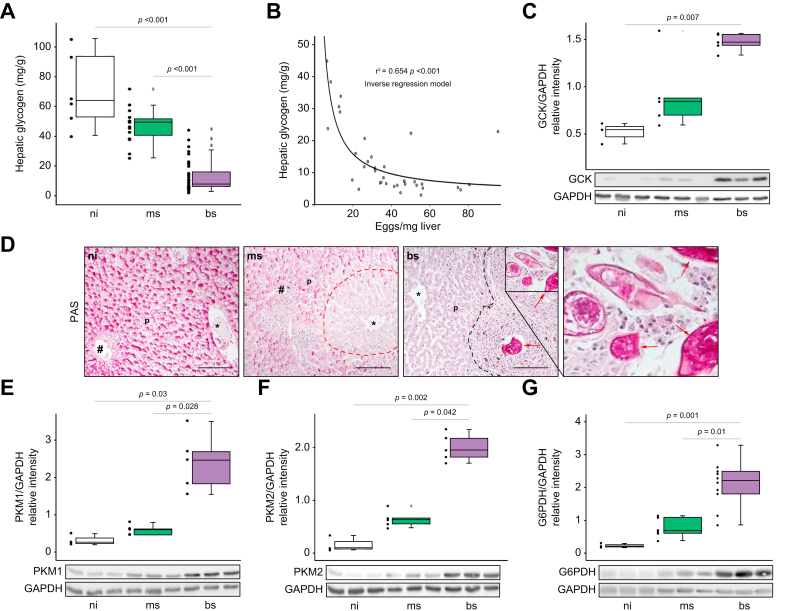
Results: The infection with S. mansoni provokes hepatic exhaustion of neutral lipids and glycogen. Furthermore, the distribution of distinct lipid species and the regulation of rate-limiting metabolic enzymes is disrupted in the liver of S. mansoni infected animals. Notably, eggs mobilize, incorporate, and store host lipids, while the associated metabolic reprogramming causes oxidative stress-induced DNA damage in hepatocytes. Administration of reactive oxygen species scavengers ameliorates these deleterious effects.
Conclusions: Our findings indicate that S. mansoni eggs completely reprogram lipid and carbohydrate metabolism via soluble factors, which results in oxidative stress-induced cell damage in the host parenchyma.
Impact and implications: The authors demonstrate that soluble egg products of the parasite S. mansoni induce hepatocellular reprogramming, causing metabolic exhaustion and a strong redox imbalance. Notably, eggs mobilize, incorporate, and store host lipids, while the metabolic reprogramming causes oxidative stress-induced DNA damage in hepatocytes, independent of the host's immune response. S. mansoni eggs take advantage of the host environment through metabolic reprogramming of hepatocytes and enterocytes. By inducing DNA damage, this neglected tropical disease might promote hepatocellular damage and thus influence international health efforts.
Keywords: DMPE, dimethyl-phosphatidylethanolamine; DNA damage; GS, glycogen synthase; GSH, reduced L-glutathione; HCC, hepatocellular carcinoma; Lipid; MALDI-MSI, matrix assisted laser desorption/ionization mass spectrometry imaging; MDA, malondialdehyde; OA, oleic acid; Oxidative stress; PAS, periodic acid-Schiff; PC, phosphatidylcholine; PDH, pyruvate dehydrogenase; PE, phosphatidylethanolamine; PLIN2, perilipin 2; Parasite; ROS, reactive oxygen species; S. japonicum, Schistosoma japonicum; S. mansoni, Schistosoma mansoni; SEA, soluble egg antigens; Schistosomiasis; TG, triglyceride; bs, bisex; flOA, fluorescently labelled OA; hRF, retention factor ∗ 100; ms, monosex; ni, non-infected.
© 2022 The Author(s).
Conflict of interest statement
B.S. and C.G.G. are consultants of TransMIT GmbH, Giessen, Germany. The other authors declare that they have no conflicts of interest. Please refer to the accompanying ICMJE disclosure forms for further details.
Figures






References
-
- WHO: Fact-Sheets Schistosomiasis 2021; Available from: https://www.who.int/news-room/fact-sheets/detail/schistosomiasis.
-
- Boissier J., Grech-Angelini S., Webster B.L., Allienne J.-F., Huyse T., Mas-Coma S., et al. Outbreak of urogenital schistosomiasis in Corsica (France): an epidemiological case study. Lancet Infect Dis. 2016;16(8):971–979. - PubMed
-
- Olveda D.U., Olveda R.M., McManus D.P., Cai P., Chau T.N., Lam A.K., et al. The chronic enteropathogenic disease schistosomiasis. Int J Inf Dis. 2014;28:193–203. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
