

The molecular structure of IFT-A and IFT-B in anterograde intraflagellar transport trains
- PMID: 36593313
- PMCID: PMC10191852
- DOI: 10.1038/s41594-022-00905-5
The molecular structure of IFT-A and IFT-B in anterograde intraflagellar transport trains
Abstract
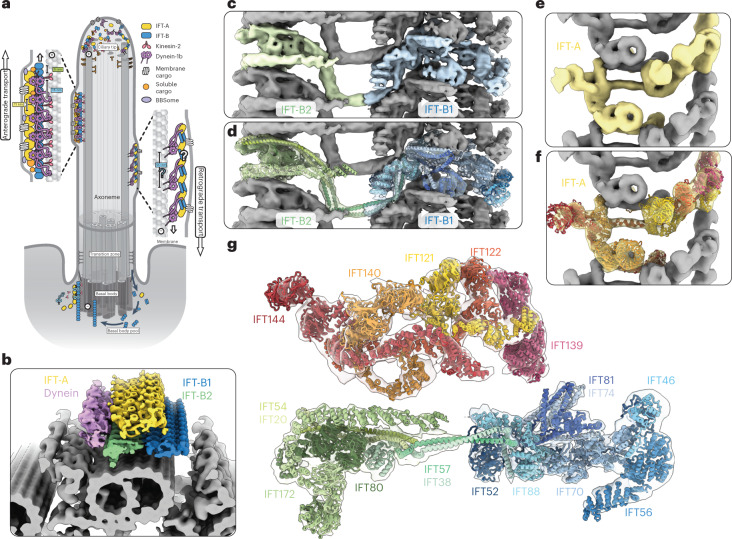
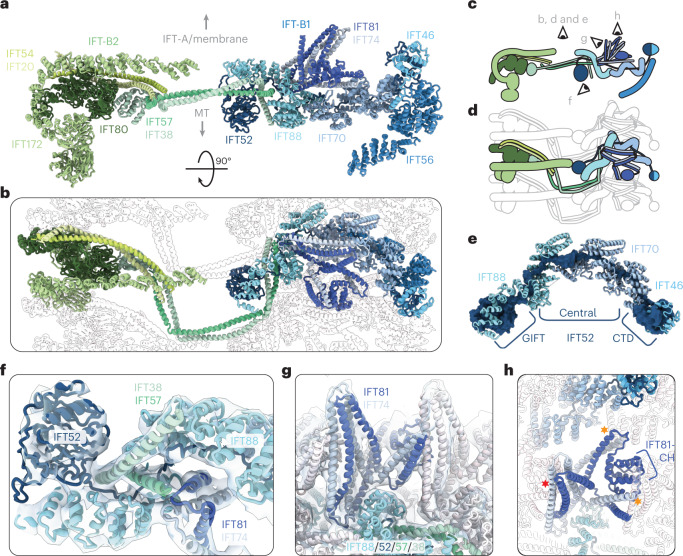
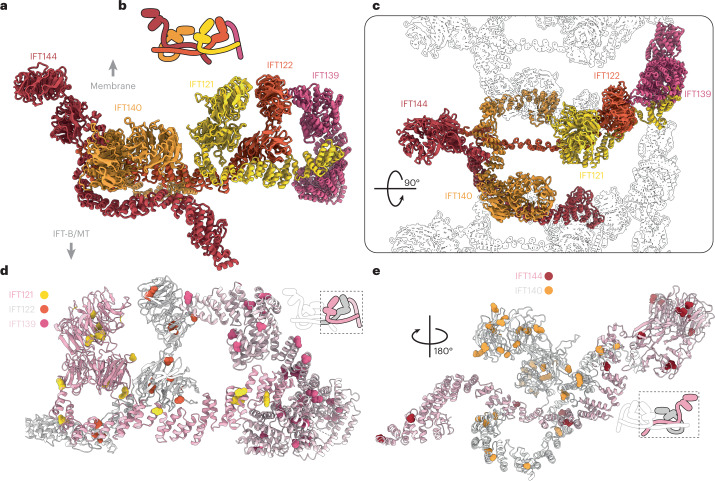
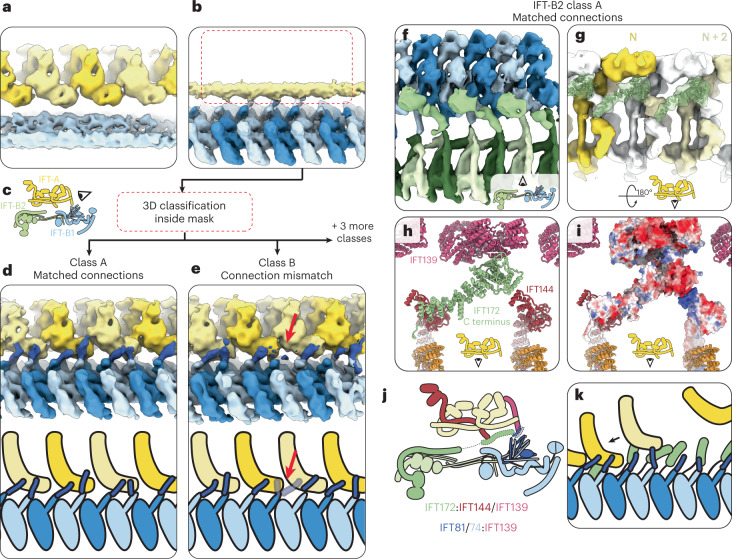
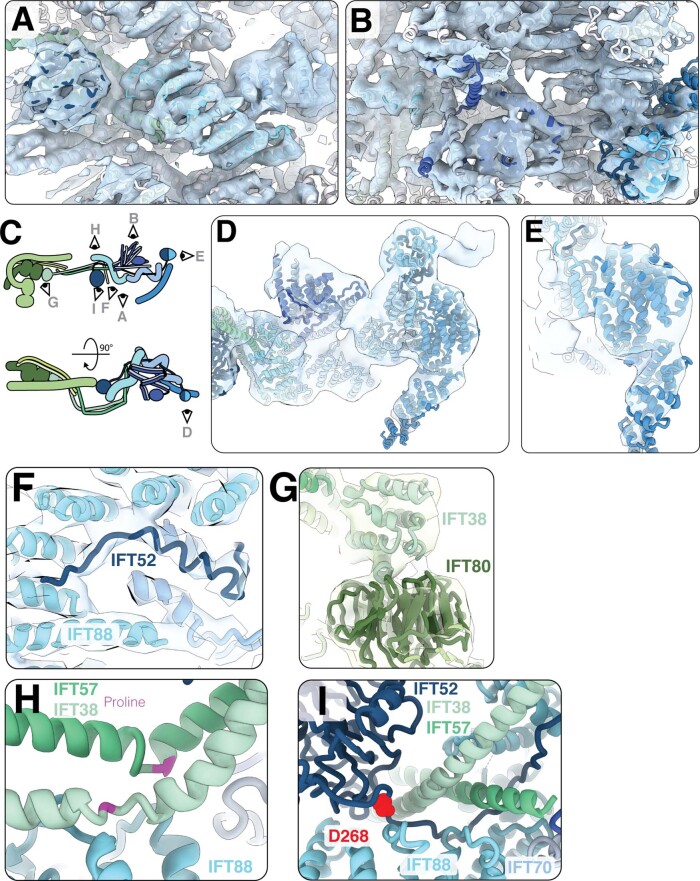
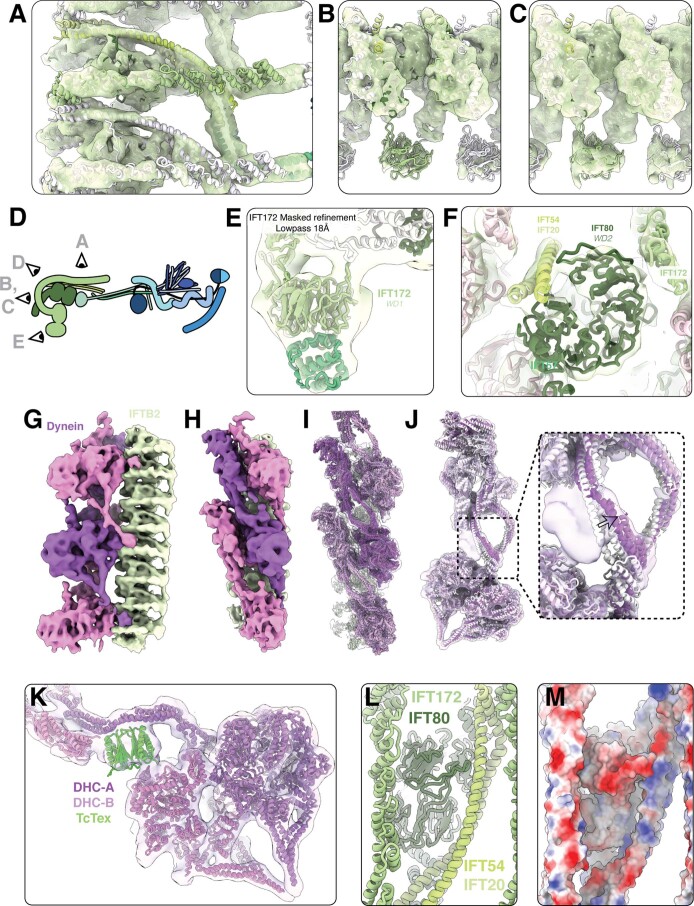
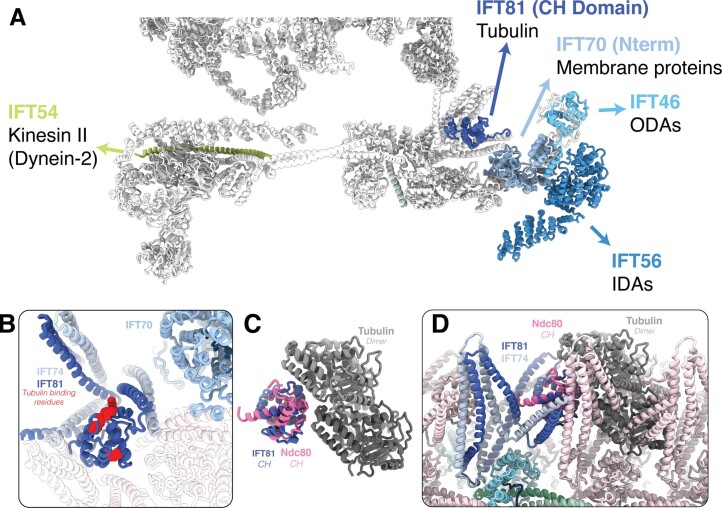
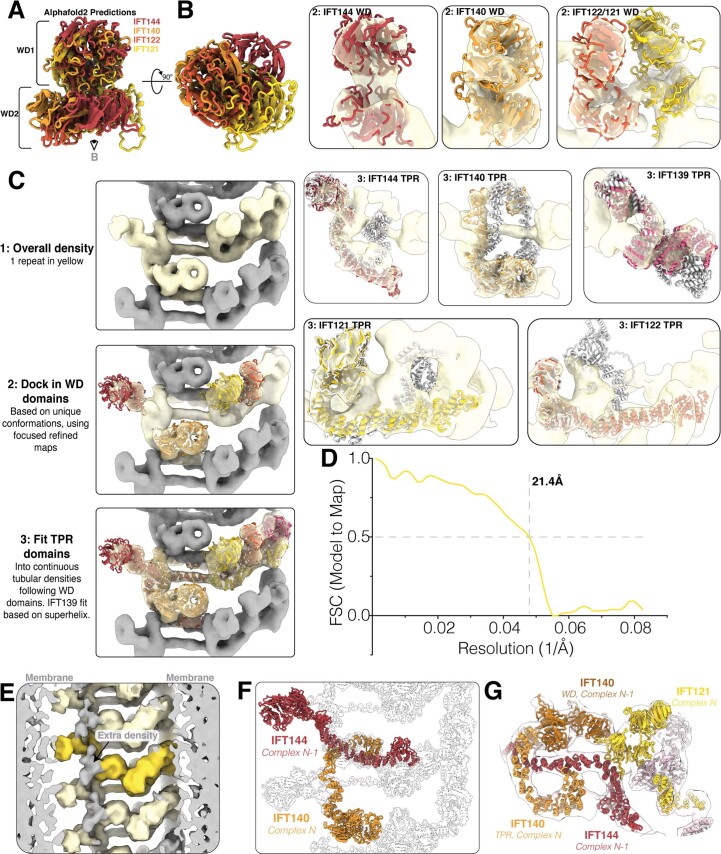
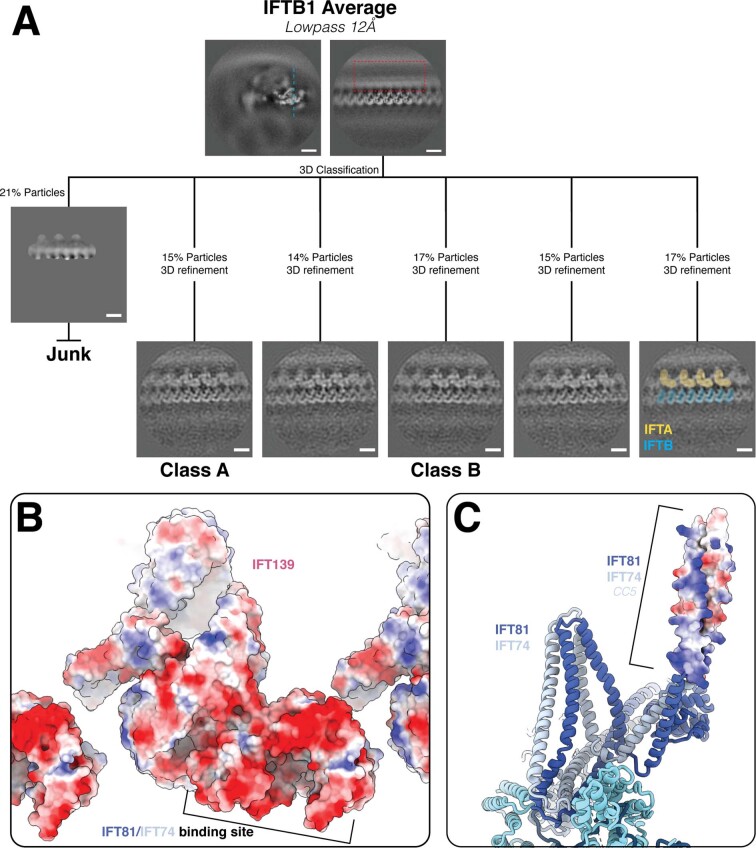
Anterograde intraflagellar transport (IFT) trains are essential for cilia assembly and maintenance. These trains are formed of 22 IFT-A and IFT-B proteins that link structural and signaling cargos to microtubule motors for import into cilia. It remains unknown how the IFT-A/-B proteins are arranged into complexes and how these complexes polymerize into functional trains. Here we use in situ cryo-electron tomography of Chlamydomonas reinhardtii cilia and AlphaFold2 protein structure predictions to generate a molecular model of the entire anterograde train. We show how the conformations of both IFT-A and IFT-B are dependent on lateral interactions with neighboring repeats, suggesting that polymerization is required to cooperatively stabilize the complexes. Following three-dimensional classification, we reveal how IFT-B extends two flexible tethers to maintain a connection with IFT-A that can withstand the mechanical stresses present in actively beating cilia. Overall, our findings provide a framework for understanding the fundamental processes that govern cilia assembly.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


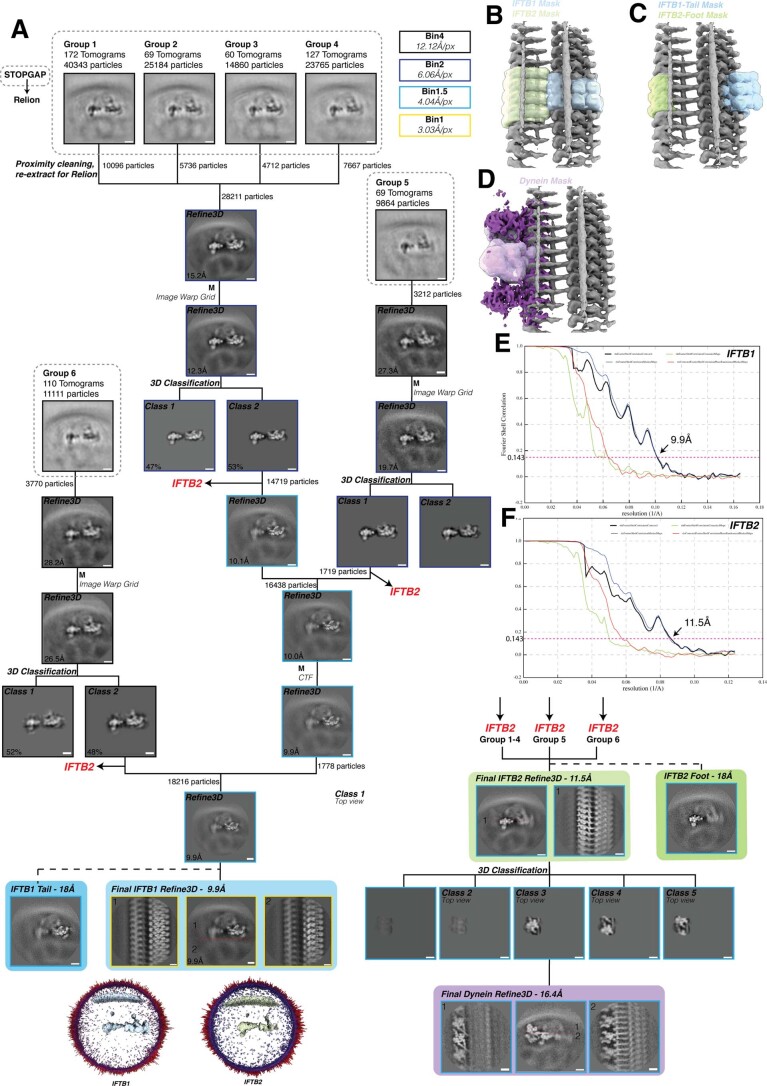
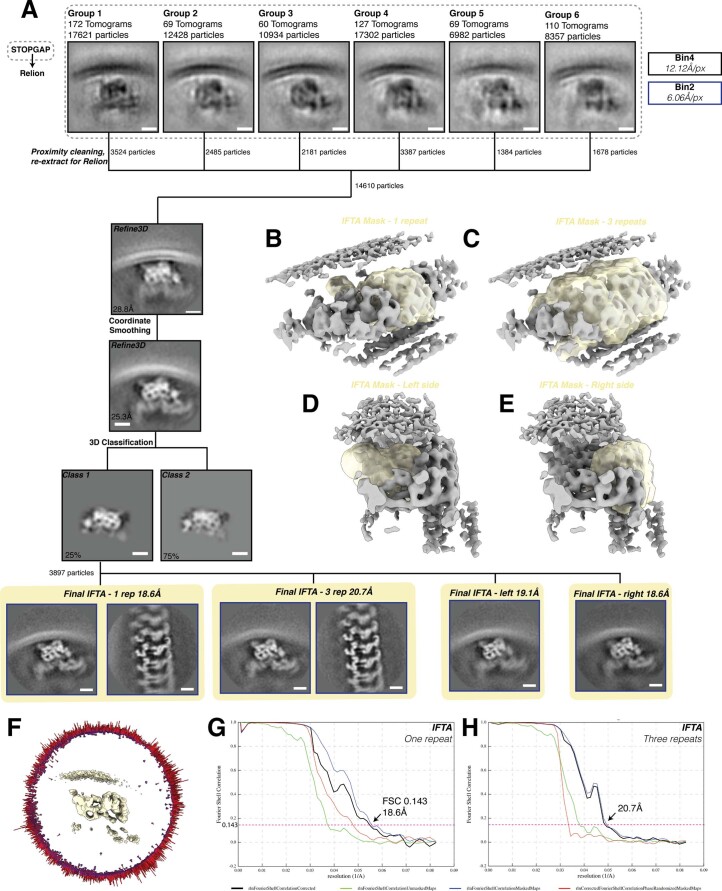
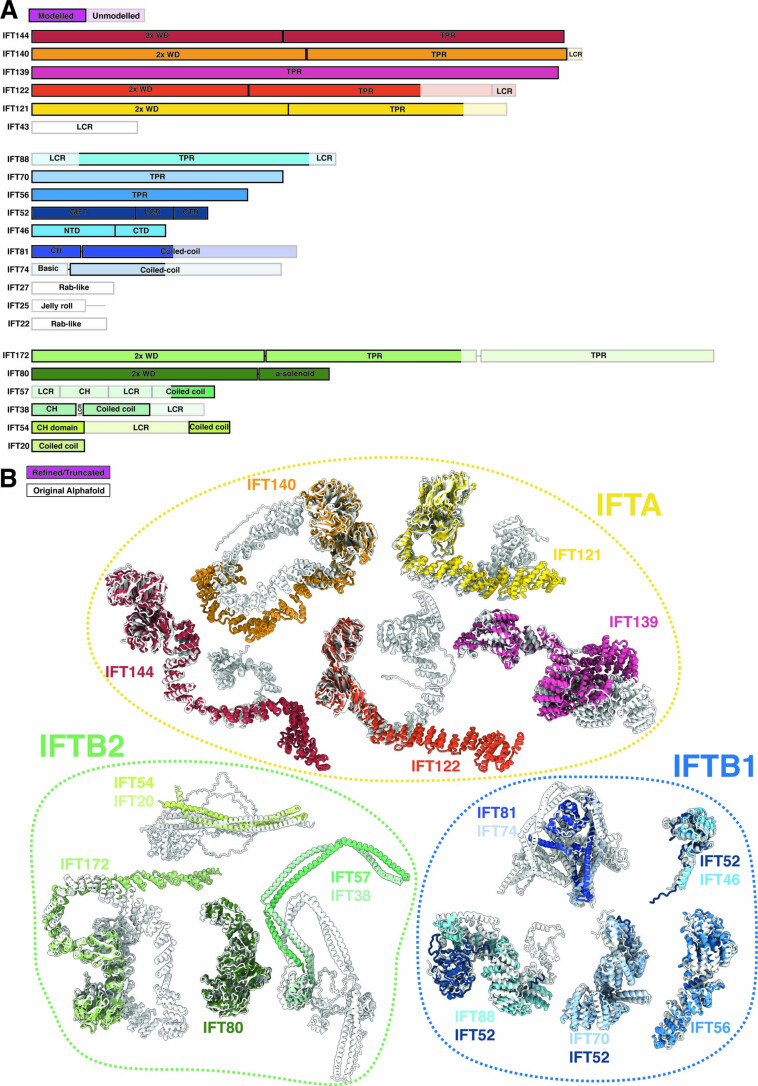
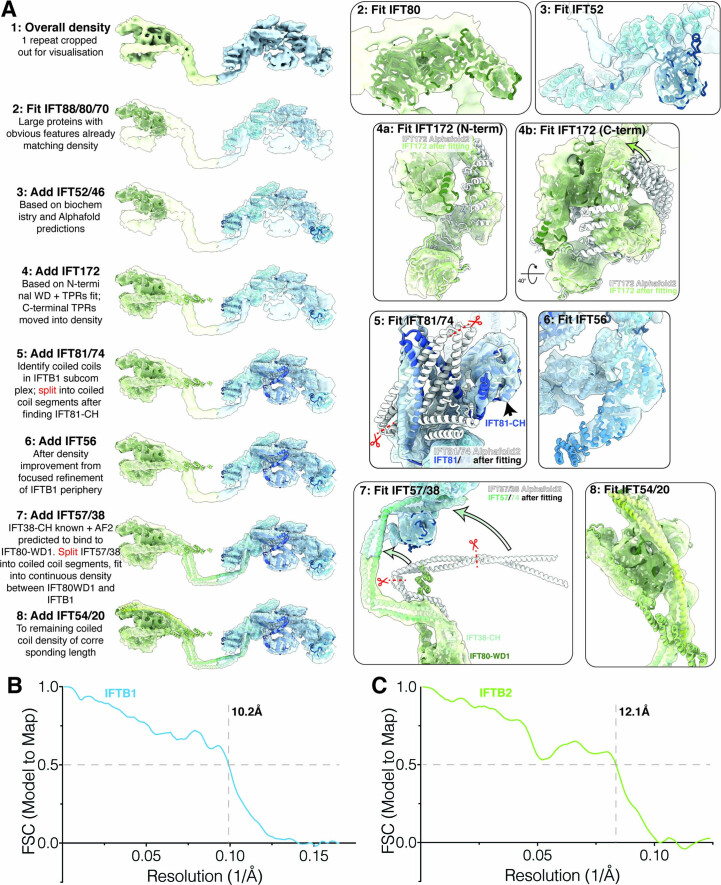
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
