

Review
doi: 10.1038/s41593-022-01222-2.
Epub 2023 Jan 2.
Insights into Alzheimer's disease from single-cell genomic approaches
Affiliations
- PMID: 36593328
- PMCID: PMC10155598
- DOI: 10.1038/s41593-022-01222-2
Item in Clipboard
Review
Insights into Alzheimer's disease from single-cell genomic approaches
Nat Neurosci.
2023 Feb.
Abstract
Alzheimer's disease (AD) is an age-related disease pathologically defined by the deposition of amyloid plaques and neurofibrillary tangles in the brain parenchyma. Single-cell profiling has shown that Alzheimer's dementia involves the complex interplay of virtually every major brain cell type. Here, we highlight cell-type-specific molecular perturbations in AD. We discuss how genomic information from single cells expands existing paradigms of AD pathogenesis and highlight new opportunities for therapeutic interventions.
© 2023. Springer Nature America, Inc.
Conflict of interest statement
Competing interests
The authors declare no competing interests related to this project.
Figures
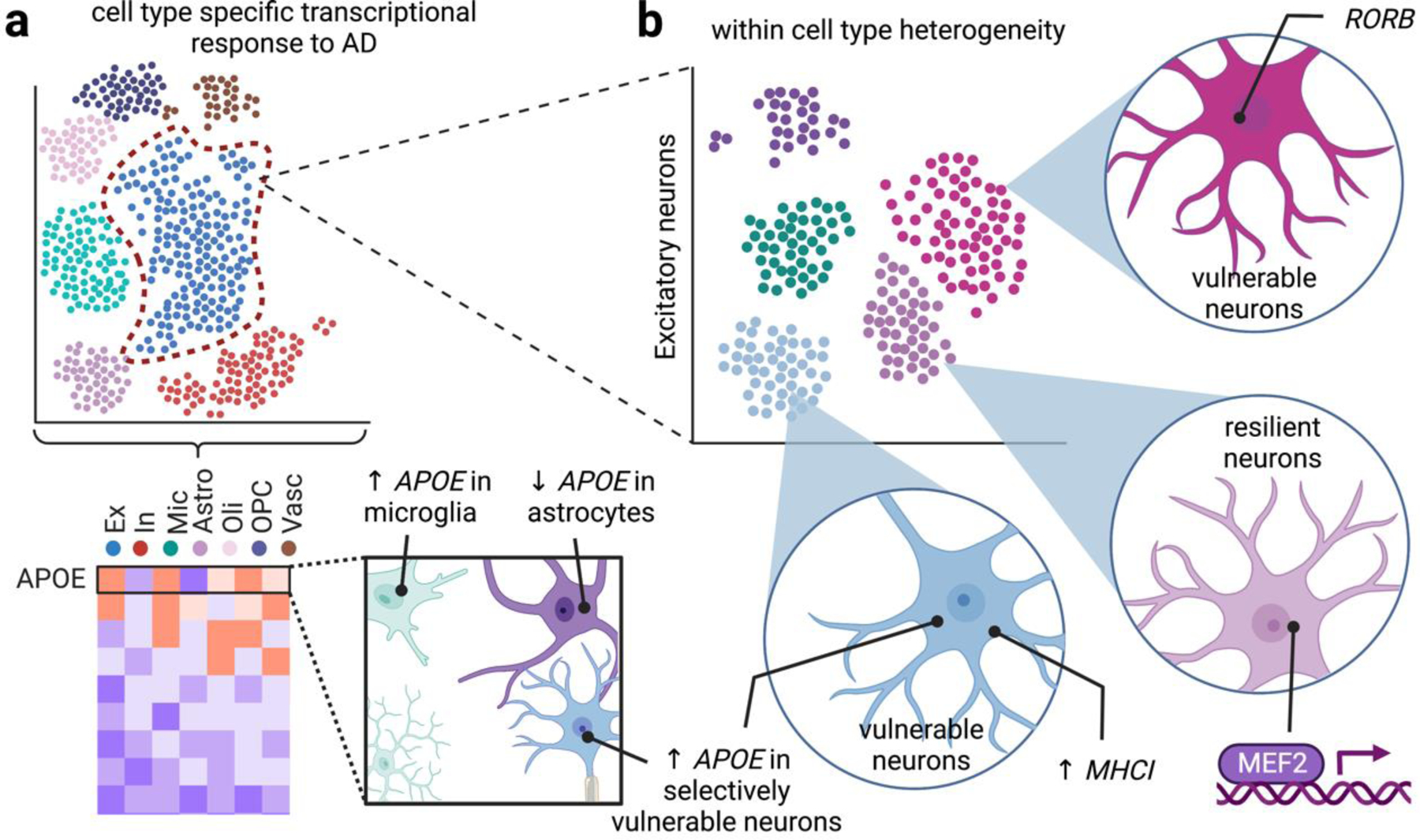
Single cell approaches highlight cell type and cell subtype specific vulnerability to disease. (a)
Cell type specific responses to disease. Bulk quantifications of gene expression report population averages, which belie changes from specific cell populations that may drive distinct pathological responses. For example, snRNA-seq revealed that APOE is downregulated in AD astrocytes but upregulated in microglia, and some neurons. (b) Cell subtype responses to disease. Bulk profiling based on cell type markers might mask within-cell type heterogeneity, such as layer-specific neurons, non-myelinating oligodendrocytes. In contrast, single cell profiling unmasks differential vulnerabilities to AD within distinct subsets of major cell types. For example, neurons selectively vulnerable to AD neurodegeneration are marked by RORB and elevated APOE/MHC-I signaling, and neurons resilient to AD pathology are enriched in MEF2.
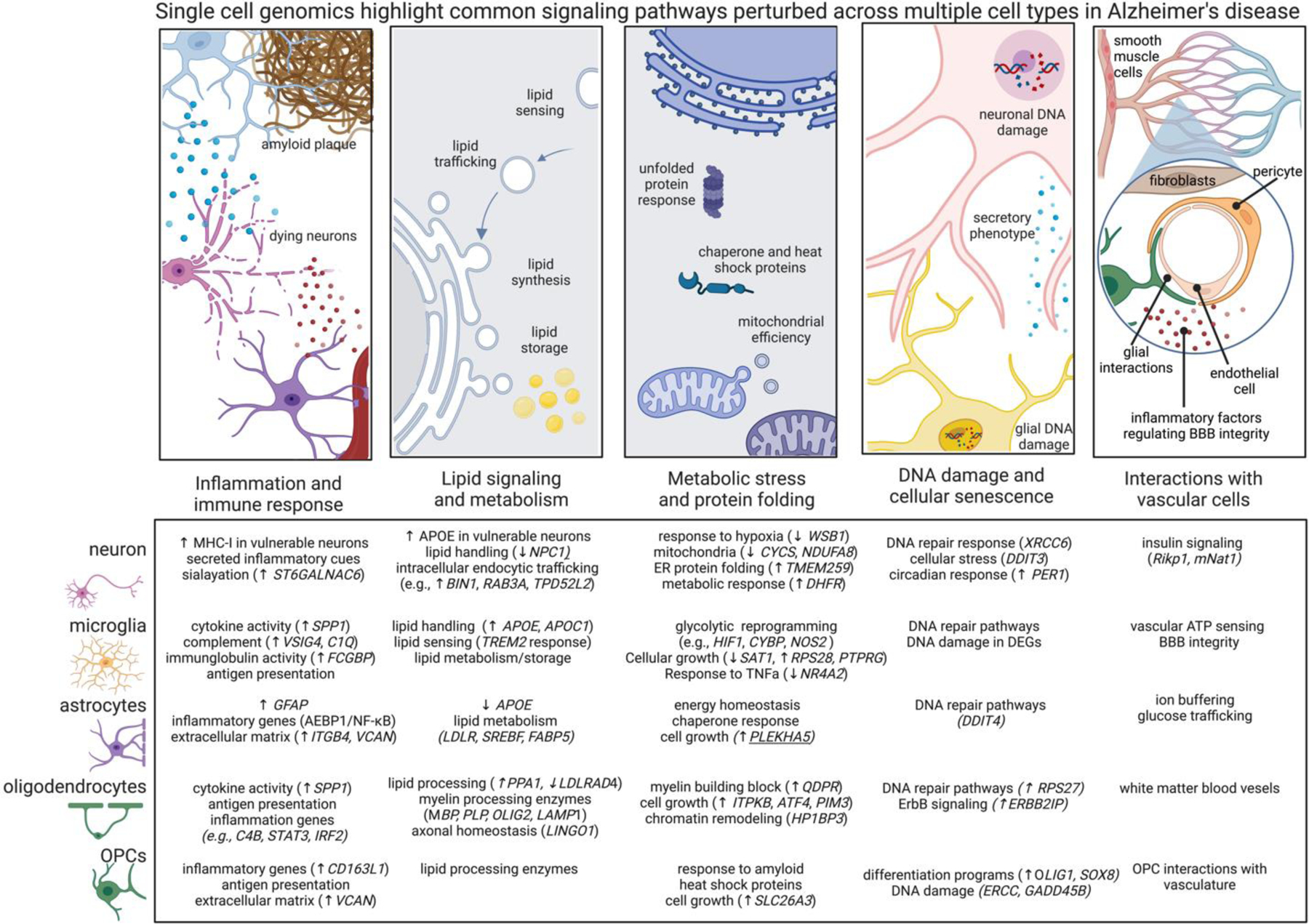
Differentially expressed genes across cell types are related to shared signaling motifs. Identifying common disrupted pathways may uncover core nodes of perturbation and lead to new therapeutic interventions related to multiple cells. Below we highlight common cellular pathways that are disrupted across multiple cell types in AD. Arrows denote transcriptional directions from prefrontal cortex (up arrow means up in AD compared to non-AD) and with a focus on genes showing concordant expression changes from other datasets and brain regions as highlighted in the text. •Immune signaling. Nearly every cell type generates immune responses in AD, including transcriptional responses related to cytokine, chemokine, and MHC signaling. MHC signaling may related to synaptic plasticity and the unfolded protein response. The low-grade AD-related inflammation in every cell type associated with AD may be associated with metabolic reprogramming. •Lipid handling. Lipid signaling is crucial for many cell functions, such as sensing and shuttling lipid species and to accommodate the dynamic remodeling of plasma membrane required for the structural plasticity of dendritic spines, microglial processes, astrocytic endfeet, and nodes of Ranvier. Perturbed lipid signaling in many brain cell types in AD, underscore the importance of lipid signaling and metabolism. •Unfolded protein response. Nearly every major cell type modulates protein misfolding pathways and integrated stress responses, and, related, mitochondrial function, highlighting energetic disruptions in AD cells. These findings suggest the milieu of the AD brain affects unfolded protein response and cellular stress even in cells not directly burdened by pathology. •DNA damage and cellular senescence. DNA damage in neurons is associated with aging and is elevated in neurodegeneration, DNA damage is essential for the expression of learning related immediate early gene expression. Many cells in AD have impaired DNA repair enzyme pathways, potentially suggesting senescent state and loss of core cellular functions. •Vascular interactions. Recent studies are beginning to profile the complex network of vascular cells in AD. Existing datasets highlight signaling pathways perturbed across multiple brain cell types relating to neurovascular coupling and BBB dysfunction in AD, including the cell-type specific secretion of inflammatory molecules known to regulate vascular cells.
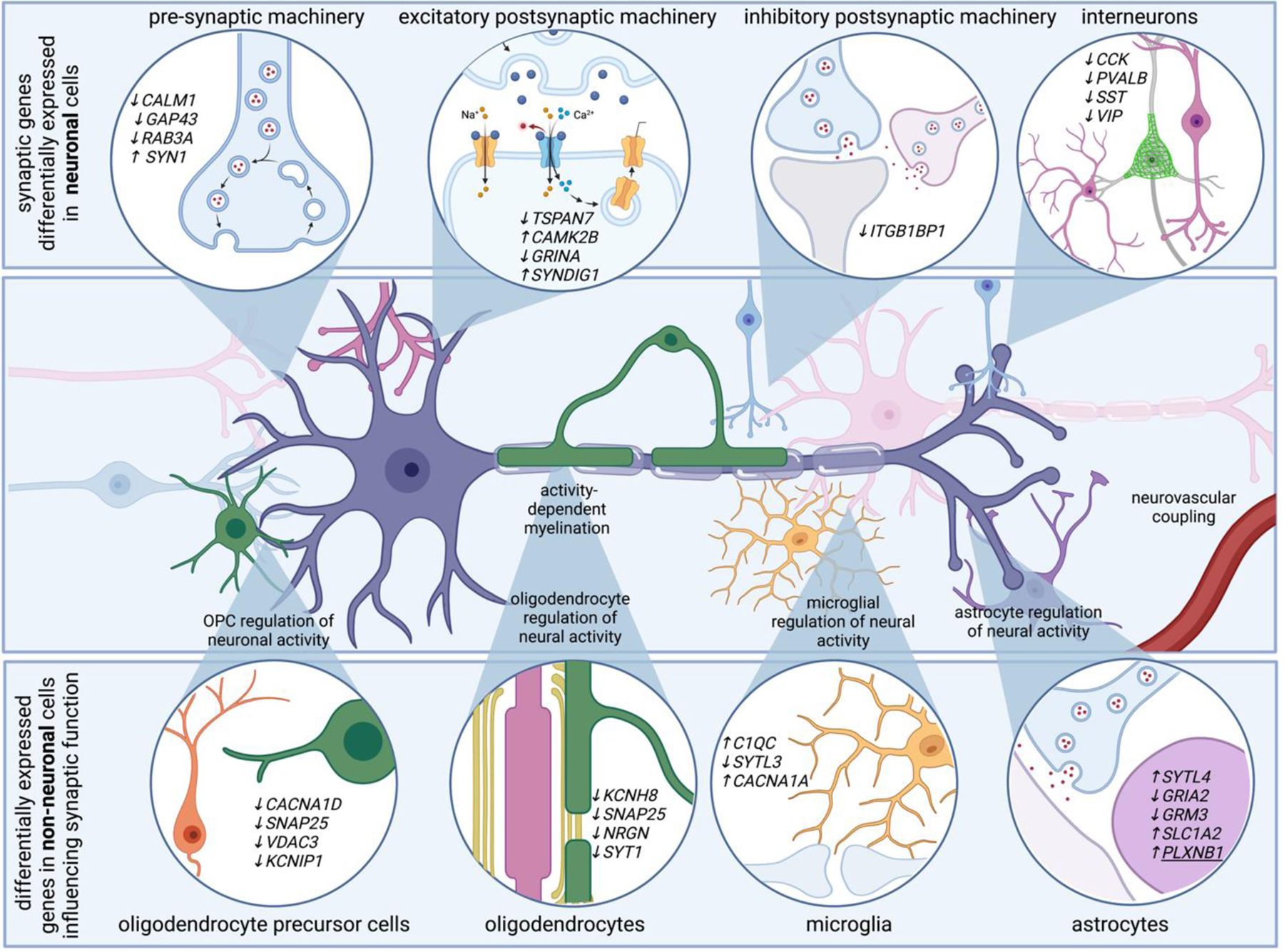
Neurons account for the vast majority of differentially expressed genes in AD. Genes related to pre-synaptic, post-synaptic, and inhibitory synaptic machinery emerge in single transcriptomes of AD neurons. For example, AD neurons upregulate SYN1, a gene that encodes synapsin 1, critical for synaptic vesicle function, and downregulate TSPAN7, which encodes a tetraspanin thought to regulate post-synaptic dendritic spine structure. Transcriptional programs associated with altered electrical properties may be associated with neuronal vulnerability to AD. Additionally, non-neuronal cells modulate genes that are involved in synaptic function. For example, genes related to synaptotagmin related genes are differentially expressed in astrocytes, oligodendrocytes, oligodendrocyte precursor cells, and microglia. Several differentially expressed genes in non-neuronal cells converge on pathways that ultimately influence neuronal function, such as genes related to synaptic pruning and activity-dependent ion channels. For example, voltage gated ion channels, which might help non-neuronal cells sense neuronal activity, are also modulated in multiple cell types. These highlight how many brain cell types are involved in sensing and regulating neural activity, and suggest neural circuit dysfunction in AD is likely the consequence of multi-cellular signaling cascades.
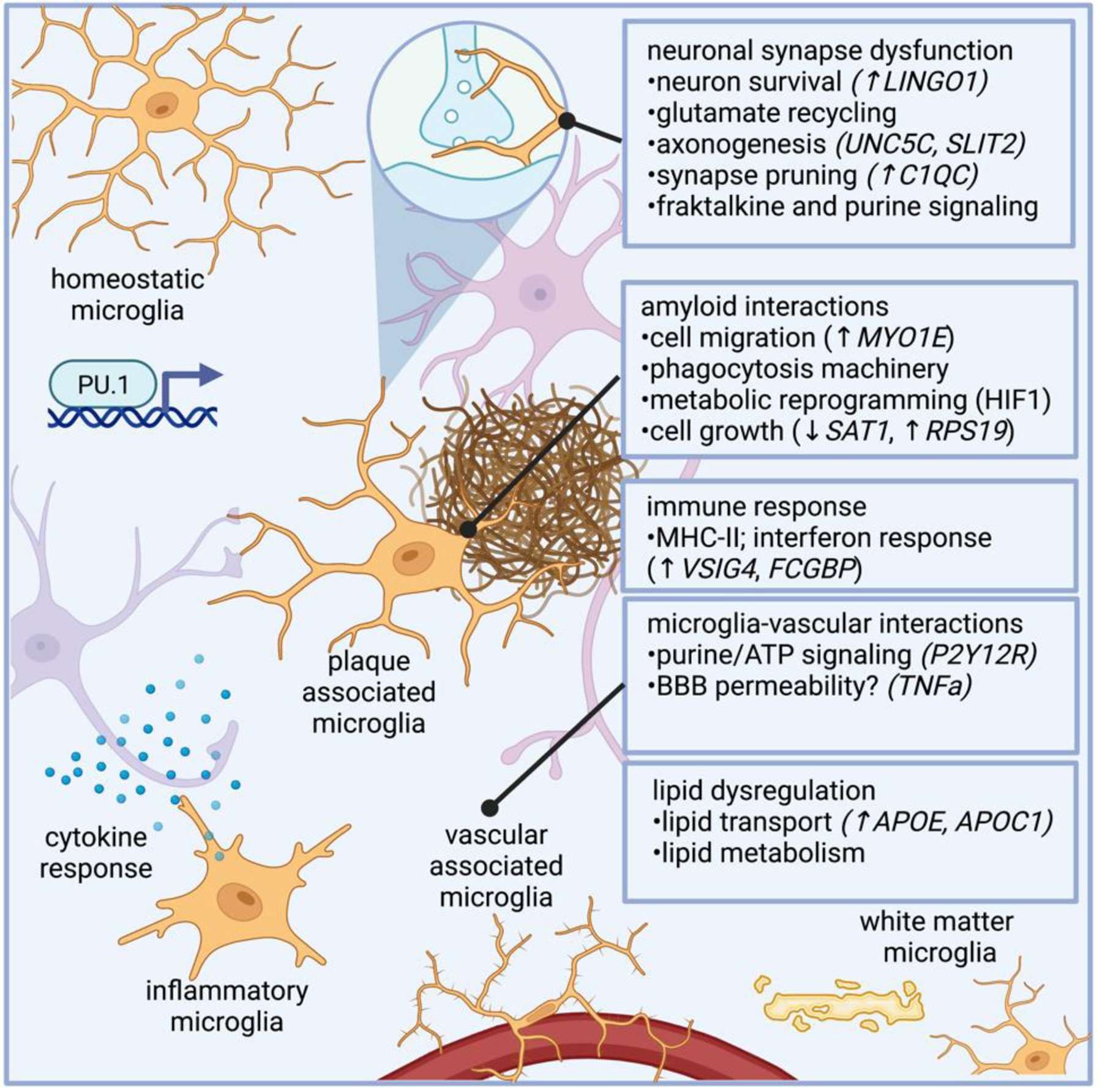
Microglia dysfunction in AD modulate genes related to synapse function, phagocytosis, and immune response. Microglia regulate genes involved in myelination, such as LINGO1, a negative regulator of myelination, as well as genes involved in axonogenesis (e.g., UNC5C and SLTI2). AD microglia also modulate complement related genes, such as C1QC, which regulate synaptic pruning. Microglial harbor properties associated with phagocytosis, and microglial response to amyloid has been well characterized, and genes in AD microglia potentially related to amyloid response include those related to microglial plaque clustering phenotypes, such as cell migration (e.g., MYO1E, which encodes a gene related to myosin), as well as genes involved in metabolic reprogramming and cell growth (e.g., SAT1, which encodes an acetyltransferase, and RPS19, a ribosomal subunit). As the innate immune cells of the brain, microglia are intimately involved in immune response, and several differentially expressed genes in AD microglia are involved in immune response, such as VSIG4 and FCGBP, genes involved in immunoglobulin response. TREM2 is a lipid receptor that is thought to govern microglia transitions to disease-associated states,. Subtypes of microglia that regulate plaques are marked by Hif1a in 5XFAD mice, which is associated with metabolic reprogramming in human AD microglia.
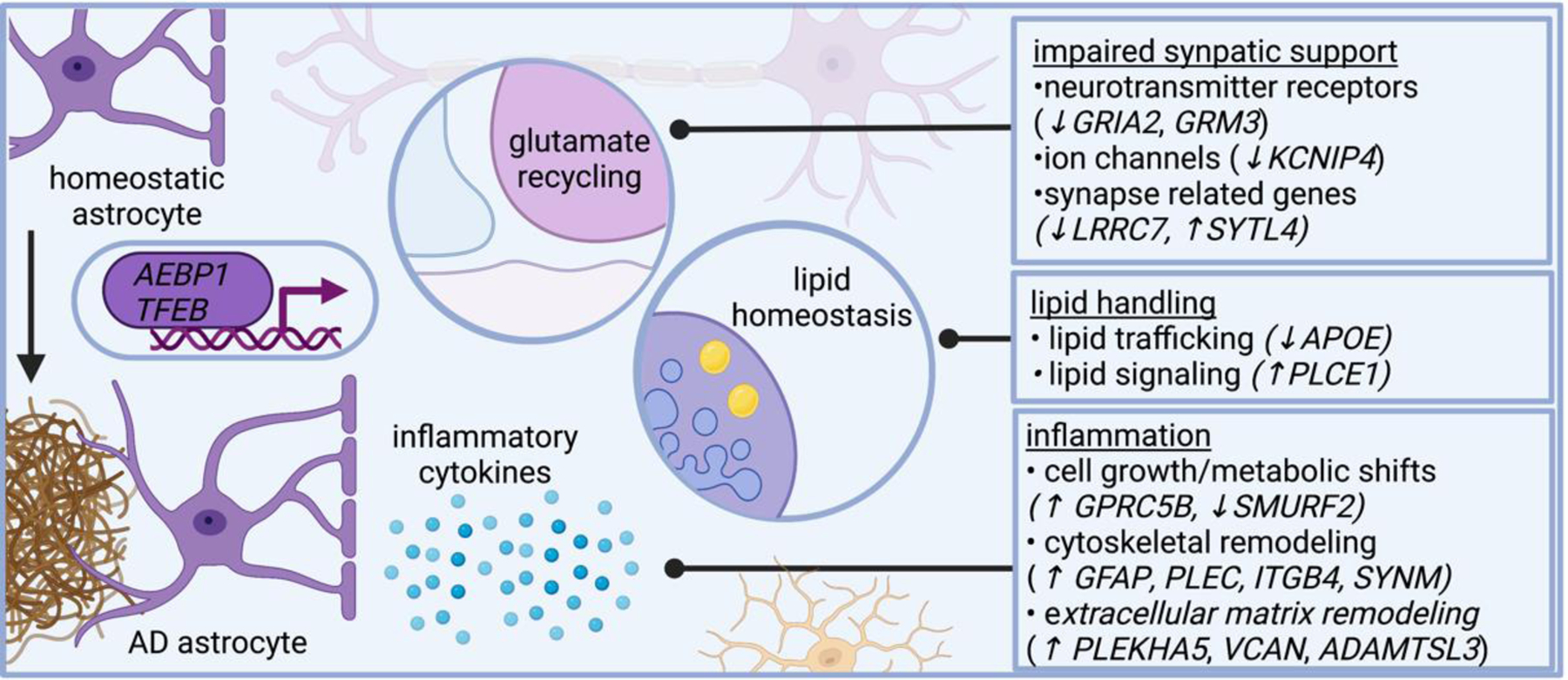
Several lines of evidence suggest astrocytes in AD become inflammatory and impair neural circuit function, including plaque-associated barriers, and modulating lipid-related signaling networks. Single cell genomics shed additional insight on these pathways and reveals astrocytes in AD modulate genes related to neurotransmitter recycling, inflammatory response, and lipid metabolism. AD dysregulates astrocytic genes involved in neurotransmitter receptors (such as GRIA2 and GRM3, which encode subunits of glutamate receptors), ion channels (such as KCNIP4, which encodes a protein that interacts with voltage-gated potassium channels), and even genes involved in synapses (such as LRRC7, which encodes a component of the post synaptic density of excitatory synapses, and SYTL4, which encodes a synaptotagmin). AD astrocytes also modulate genes involved in lipid metabolism, including APOE and PLCE1, which encodes a phospholipase. Several astrocytic genes differentially expressed in AD relate to cytoskeletal remodeling, including GFAP (which encodes an intermediate filament), PLEC (which encodes plectin, a protein that interacts with intermediate filaments), SYNM (which encodes another intermediate filament), and ITGB4 (which encodes an integrin). AD astrocytes modify genes involved in cell growth, such as SMURF2 (a member of the SMAD family important for cell growth). Collectively, these transcriptional changes highlight signaling pathways altered in AD astrocytes.
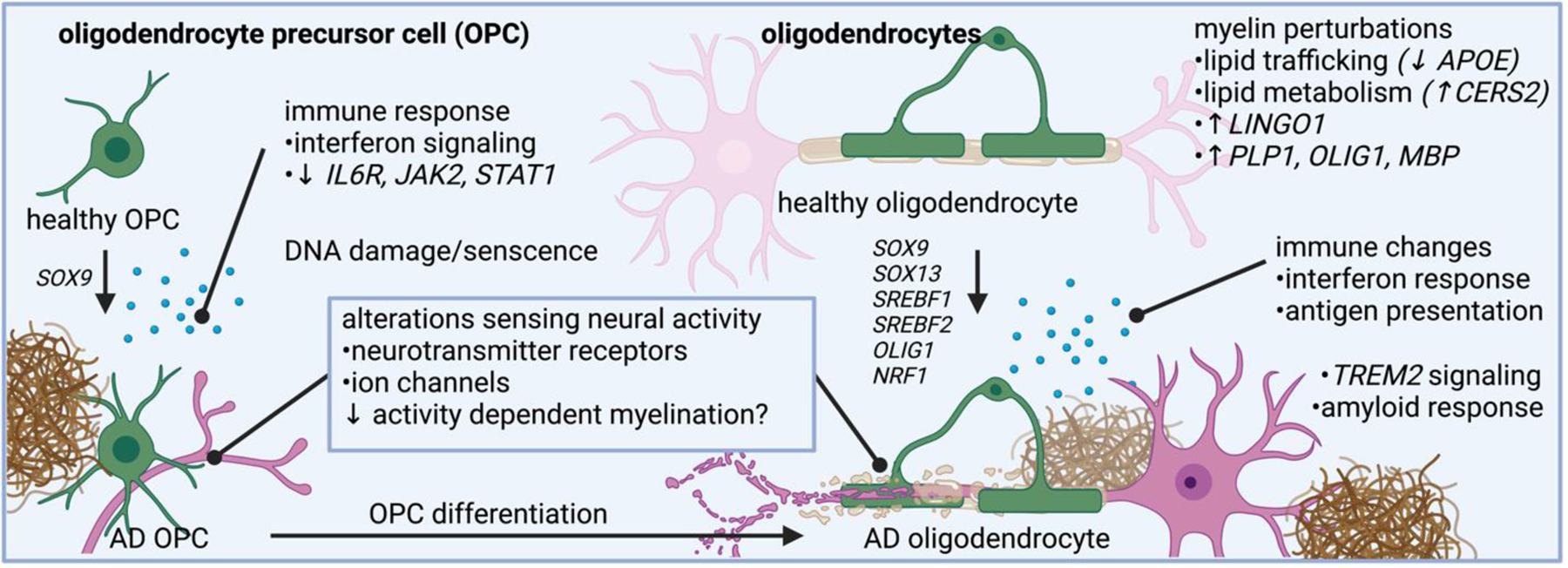
Oligodendrocytes in AD have altered pathways related to myelin synthesis, lipid trafficking, lipid metabolism, and immune related changes. Oligodendrocyte precursor cells also changes expression of genes related to neurotransmitter sensing and immune response.
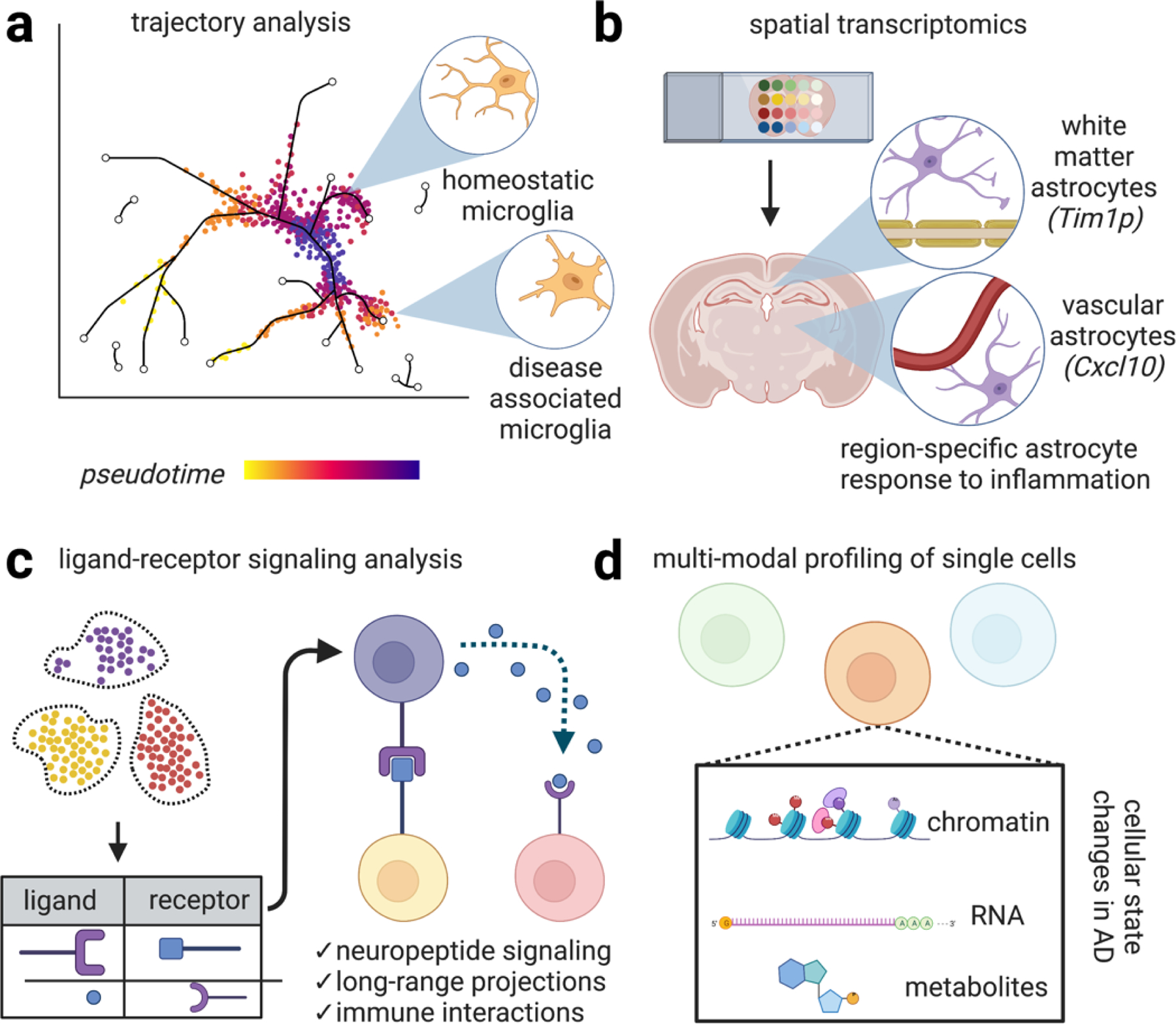
Emerging methods in single cell profiling will enhance our understanding of the distinct cellular signaling networks perturbed in AD. (a) Genetic analysis of single cells potentially enables the construction of dynamical cellular models according to “pseudotime,” a quantitative measure of biological progression through a cellular process. Applying these models to AD potentially enables the trajectory of distinct cell types adopting new transcriptional states relating to disease progression, progression of microglia from homeostatic states to disease-associated states. (b) Spatial transcriptomics is an umbrella term referring to techniques that combine mRNA readouts with spatial information. For example, one study revealed reactive subsets of astrocytes occupy distinct anatomical locations, such one astrocyte population lining white matter tracts that express the matrix metalloprotease inhibitor Timp1, which has been shown to be involved in amyloid response and has been shown to drive oligodendrocyte production and myelination and another astrocyte population marked by the cytokine Cxcl10+ adjacent to blood vessels. Preserving spatial information while performing single cell profiling will likely further enhance our understanding of the molecular mechanisms driving AD. (c) Ligand-receptor signaling involves predicting signaling interactions based on ligand/receptor databases. This analysis has revealed, for example, dense peptidergic intracortical signaling networks. Expanding our understanding of cellular physical and signaling interactions between brain cells with enhanced methods of examining these interaction networks will undoubtedly yield important insight into the progression of signaling networks in AD. (d) Multi-modal profiling of single cells to simultaneously chromatin, RNA, and potentially metabolites are emerging methods. These studies have revealed additional levels of regulation in distinct cell types in AD. Combined with enhanced sequencing depth, these methods will generate a richer portrait of cellular function in neurodegeneration.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
