

TH17 cell heterogeneity and its role in tissue inflammation
- PMID: 36596896
- PMCID: PMC10795475
- DOI: 10.1038/s41590-022-01387-9
TH17 cell heterogeneity and its role in tissue inflammation
Abstract
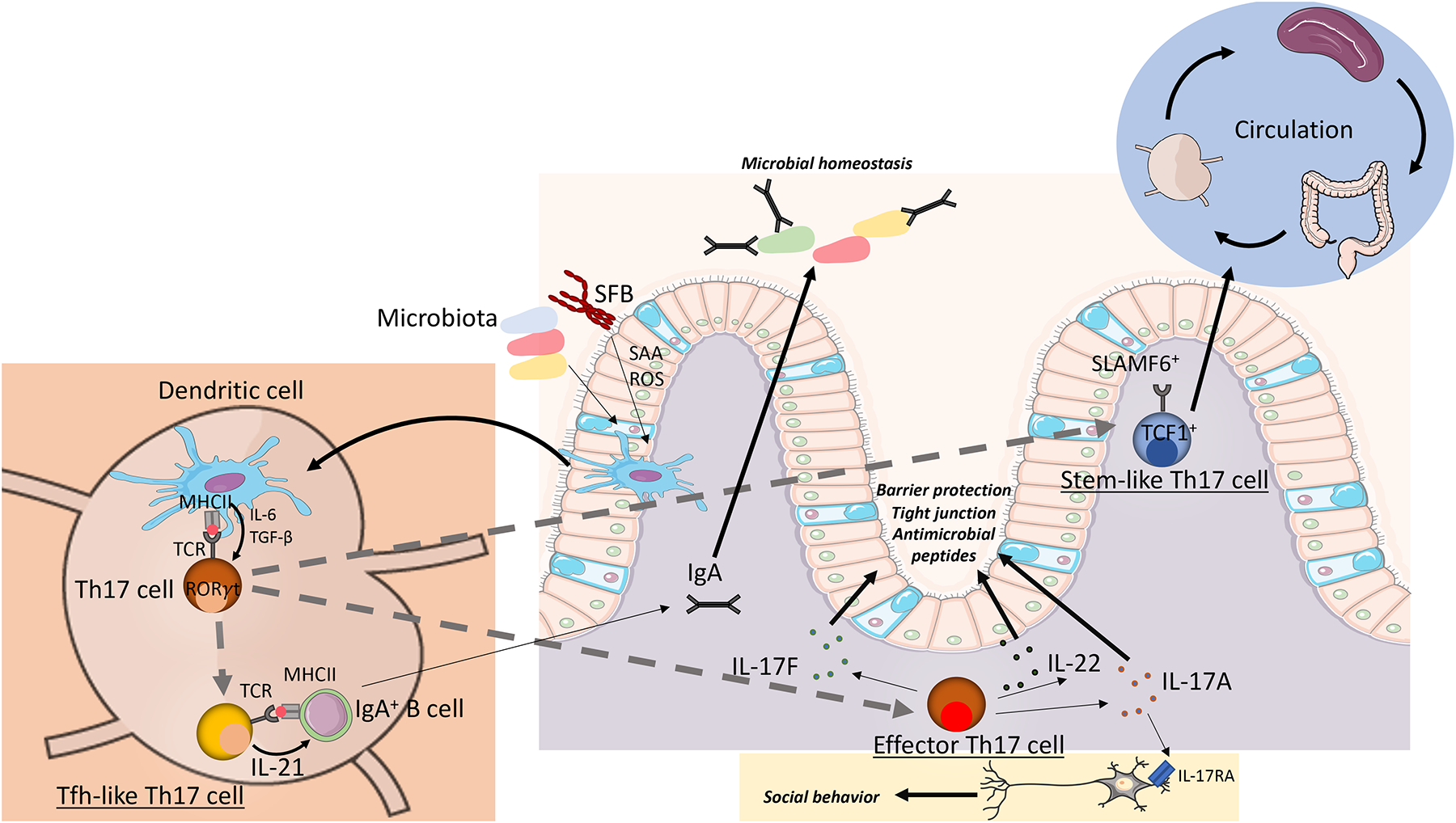
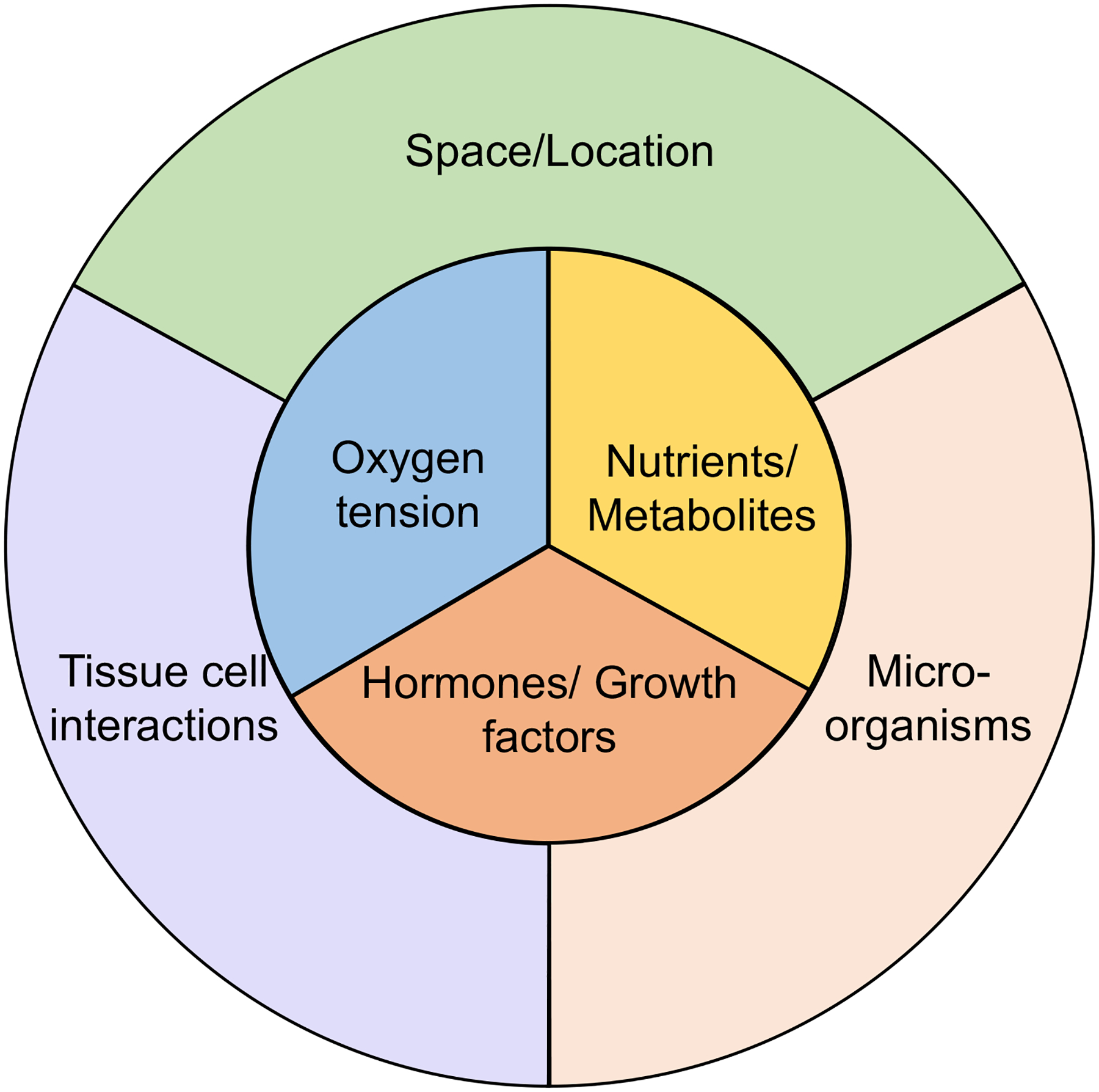
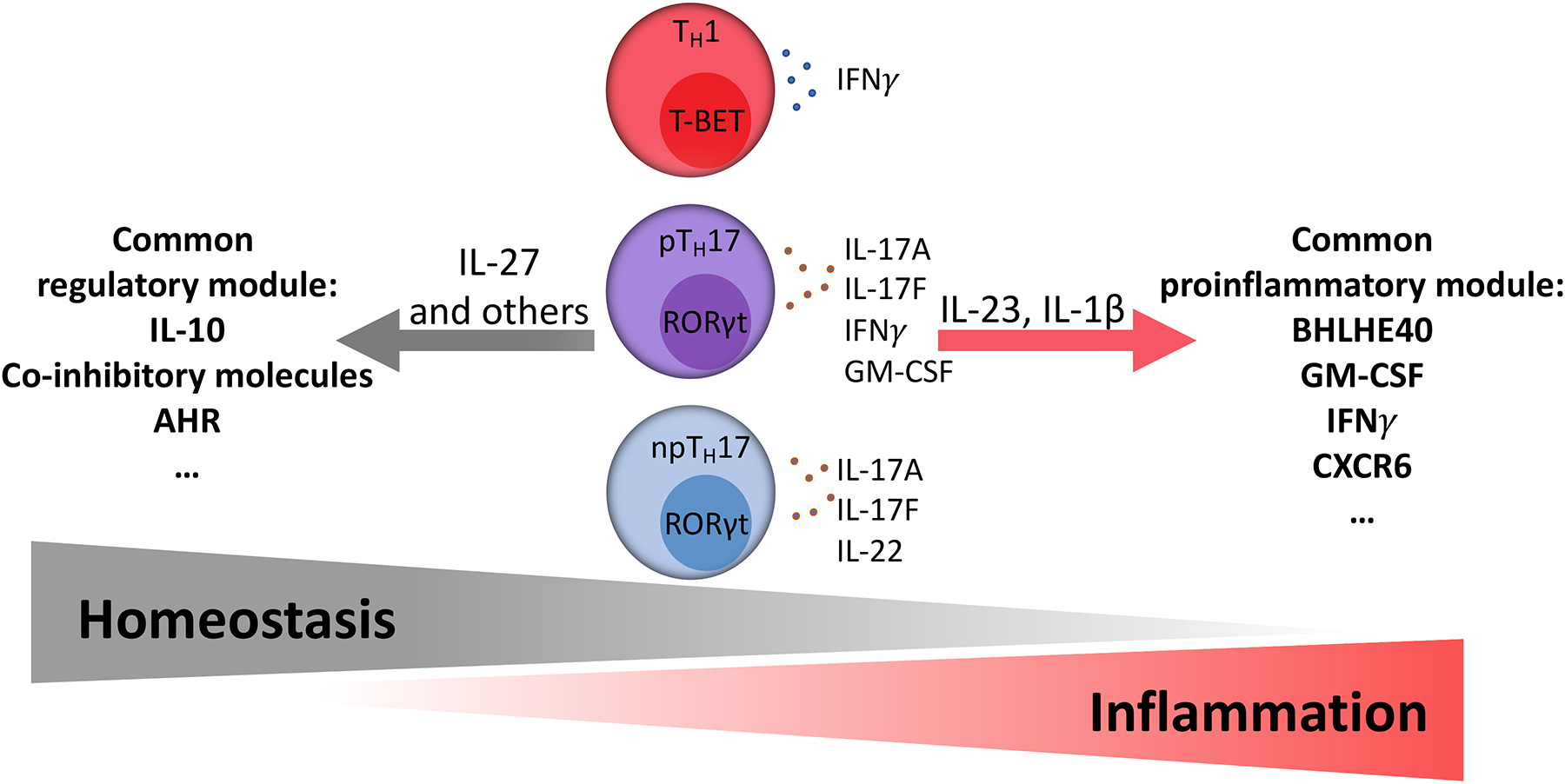
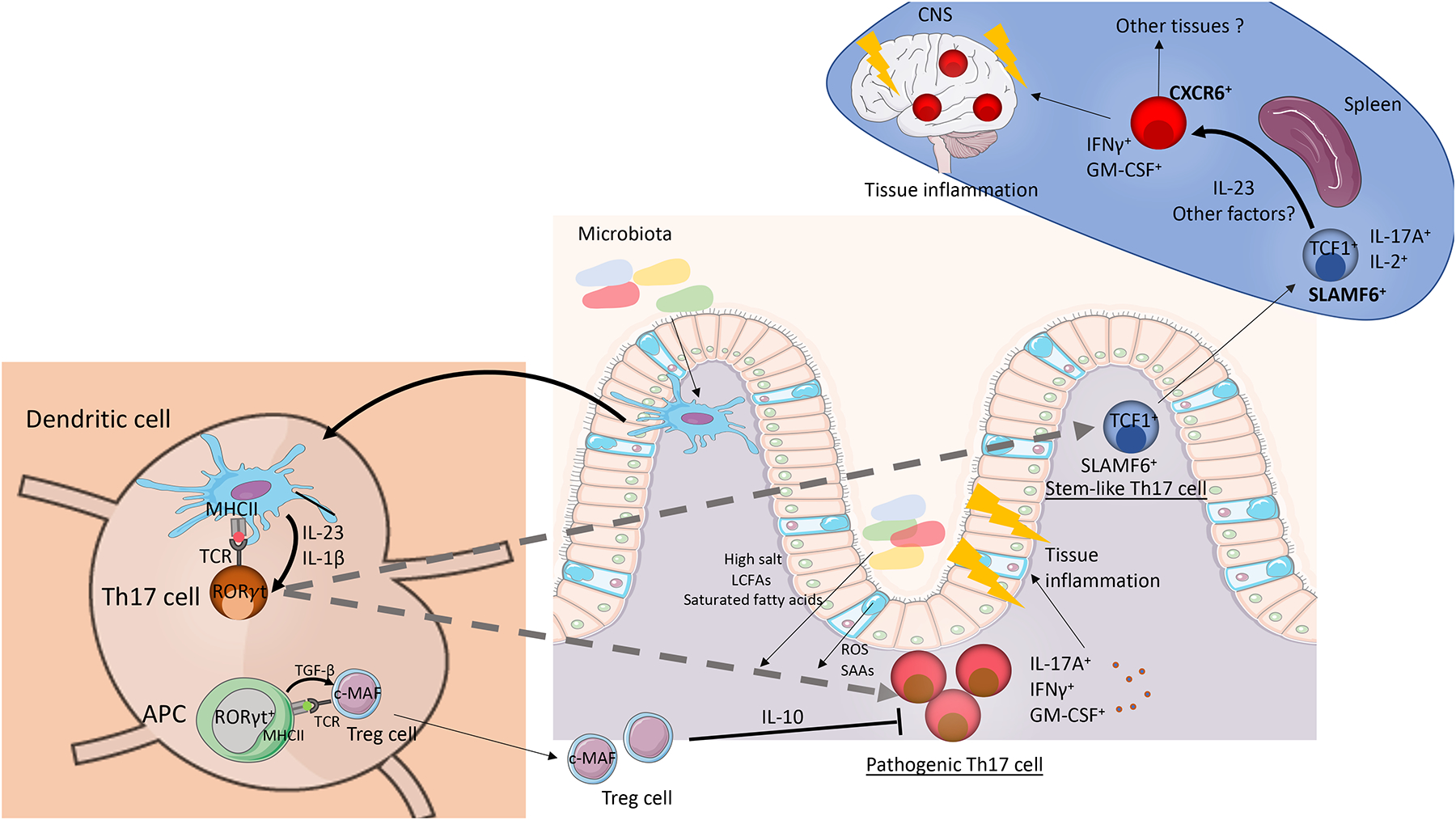
Since their discovery almost two decades ago, interleukin-17-producing CD4+ T cells (TH17 cells) have been implicated in the pathogenesis of multiple autoimmune and inflammatory disorders. In addition, TH17 cells have been found to play an important role in tissue homeostasis, especially in the intestinal mucosa. Recently, the use of single-cell technologies, along with fate mapping and various mutant mouse models, has led to substantial progress in the understanding of TH17 cell heterogeneity in tissues and of TH17 cell plasticity leading to alternative T cell states and differing functions. In this Review, we discuss the heterogeneity of TH17 cells and the role of this heterogeneity in diverse functions of TH17 cells from homeostasis to tissue inflammation. In addition, we discuss TH17 cell plasticity and its incorporation into the current understanding of T cell subsets and alternative views on the role of TH17 cells in autoimmune and inflammatory diseases.
© 2023. Springer Nature America, Inc.
Conflict of interest statement
Competing interests
V.K.K. is cofounder of Celsius Therapeutics, Tizona Therapeutics, Larkspur Biosciences and Bicara Therapeutics. His interests are reviewed and managed by the Brigham and Women’s Hospital and Partners Healthcare in accordance with their conflict of interest policies. D.R.L. is co-founder of Vedanta Biosciences and Immunai, on advisory boards of Chemocentryx and Imidomics, and on the board of directors of Pfizer, Inc. All other authors declare no competing interests.
Figures
References
-
- Mosmann TR and Coffman RL, TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol, 1989. 7: p. 145–73. - PubMed
-
- Krakowski M and Owens T, Interferon-gamma confers resistance to experimental allergic encephalomyelitis. Eur J Immunol, 1996. 26(7): p. 1641–6. - PubMed
-
- Tran EH, Prince EN, and Owens T, IFN-gamma shapes immune invasion of the central nervous system via regulation of chemokines. J Immunol, 2000. 164(5): p. 2759–68. - PubMed
-
- Gran B, et al., IL-12p35-deficient mice are susceptible to experimental autoimmune encephalomyelitis: evidence for redundancy in the IL-12 system in the induction of central nervous system autoimmune demyelination. J Immunol, 2002. 169(12): p. 7104–10. - PubMed
-
- Zhang GX, et al., Induction of experimental autoimmune encephalomyelitis in IL-12 receptor-beta 2-deficient mice: IL-12 responsiveness is not required in the pathogenesis of inflammatory demyelination in the central nervous system. J Immunol, 2003. 170(4): p. 2153–60. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
