

Structures and mechanisms of tRNA methylation by METTL1-WDR4
- PMID: 36599982
- PMCID: PMC9930641
- DOI: 10.1038/s41586-022-05565-5
Structures and mechanisms of tRNA methylation by METTL1-WDR4
Abstract
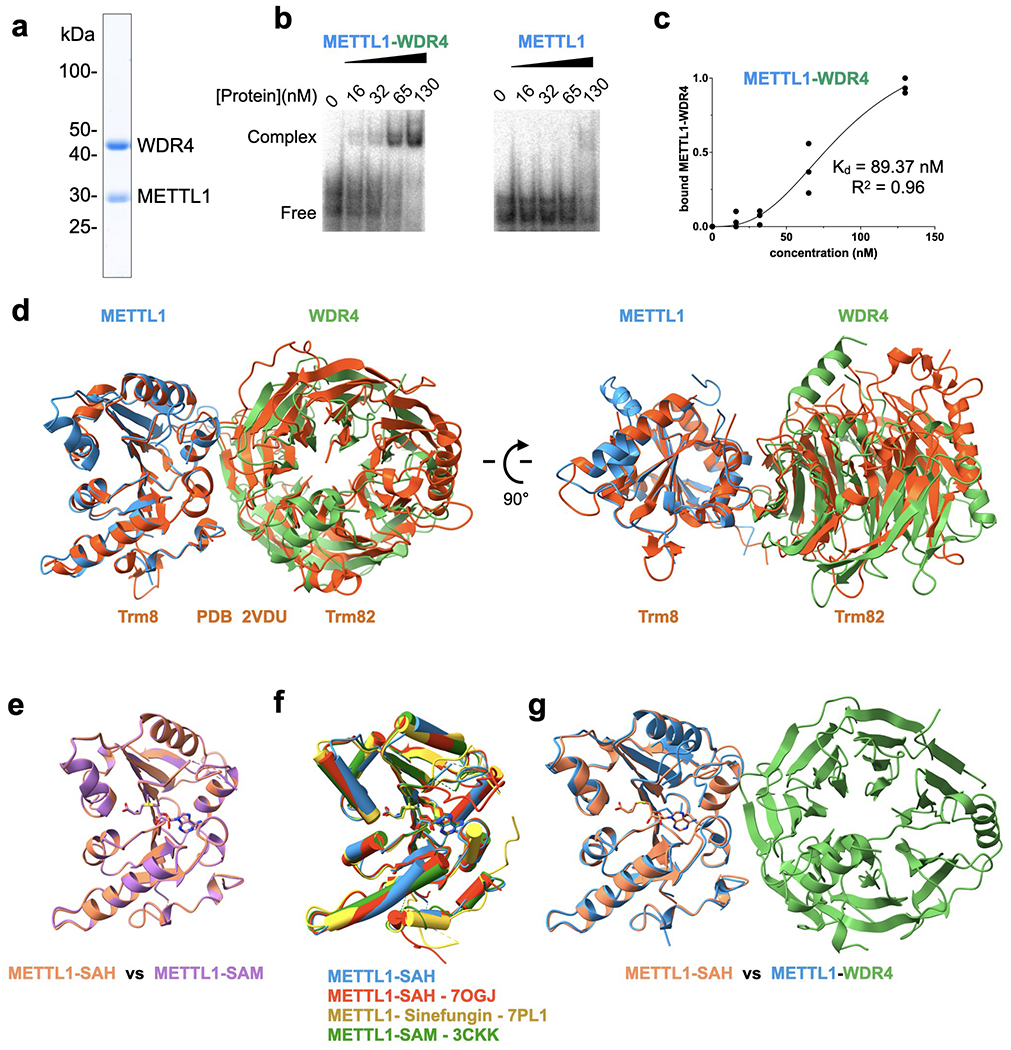
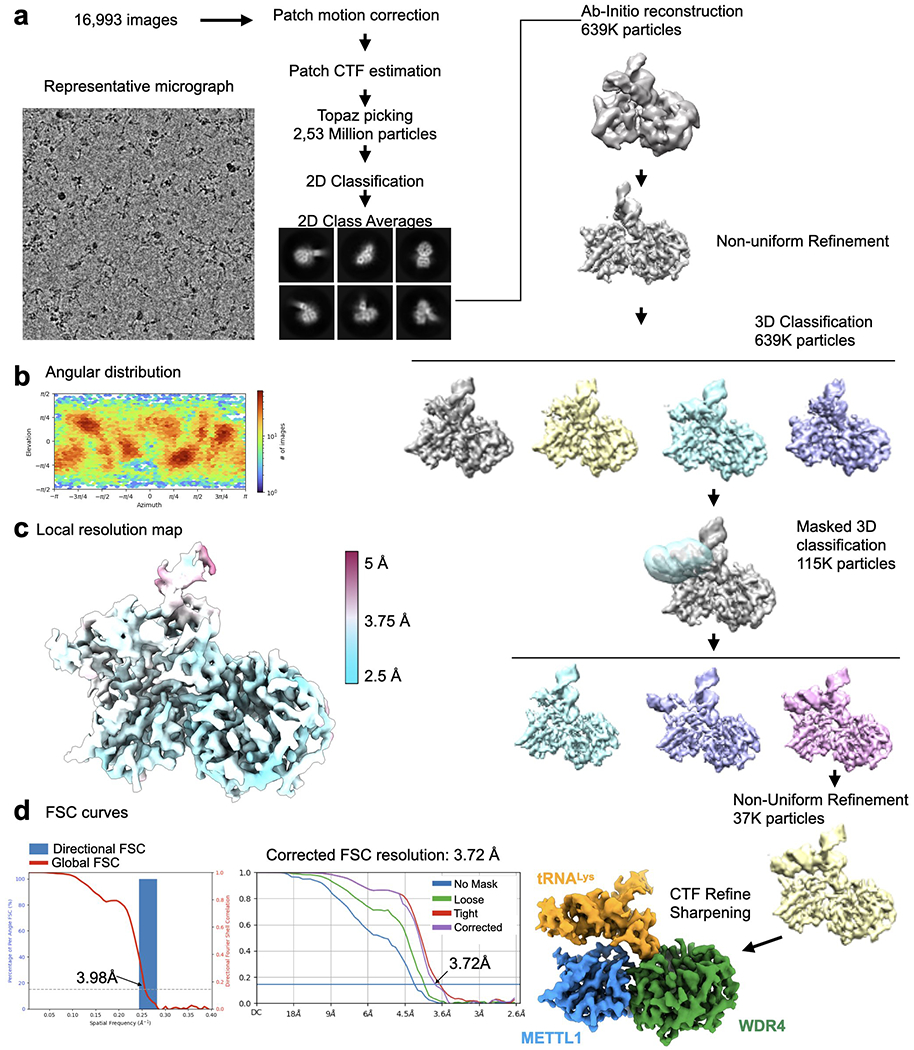
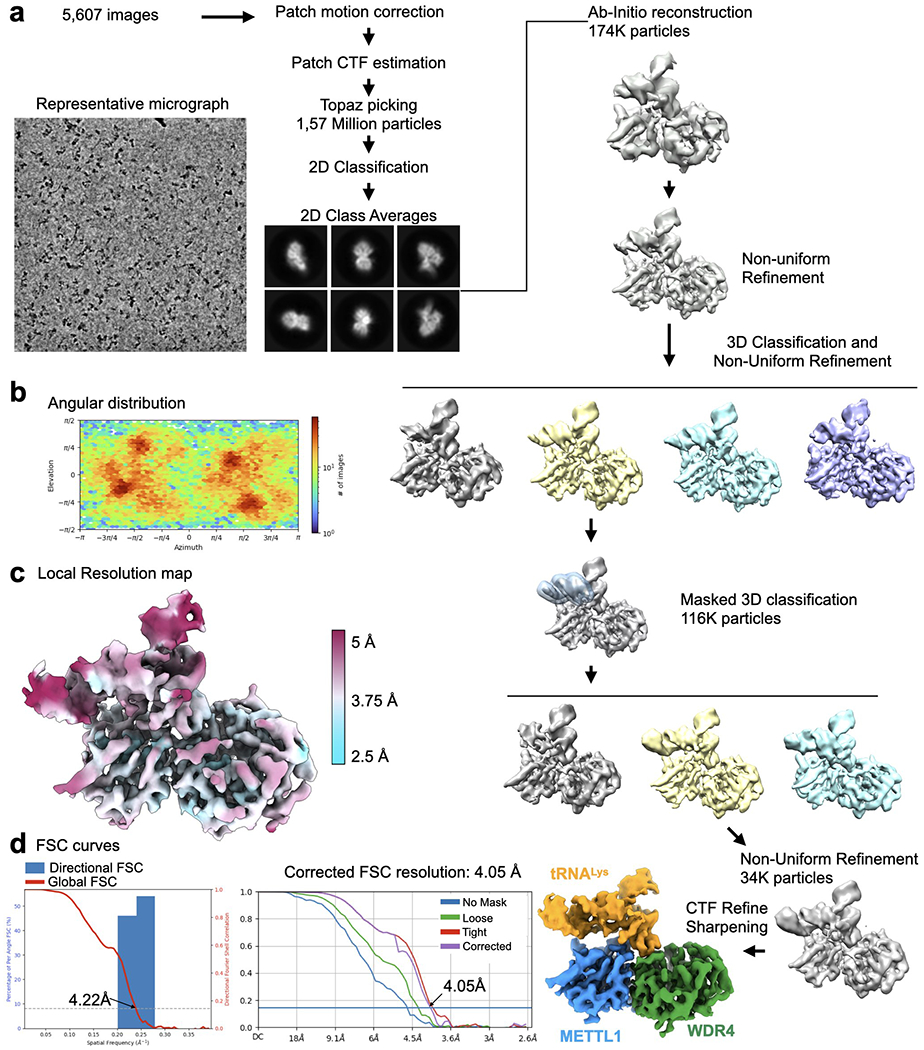
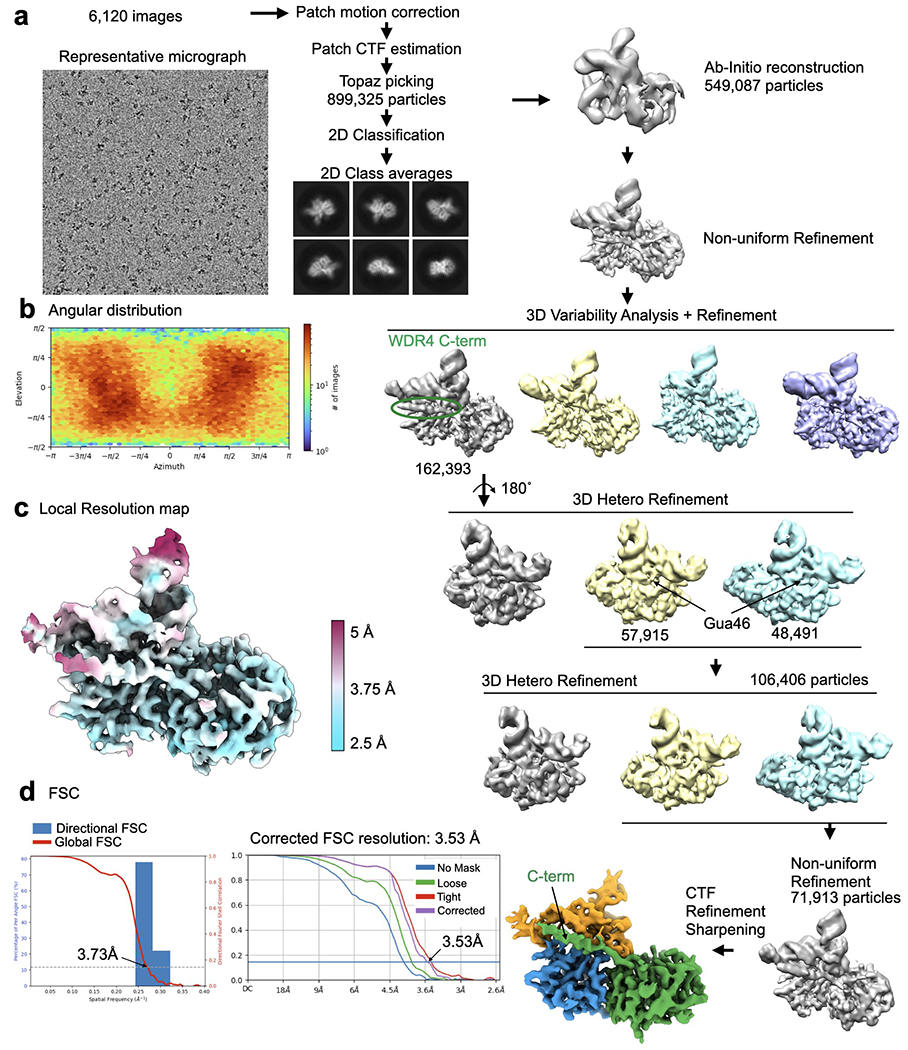
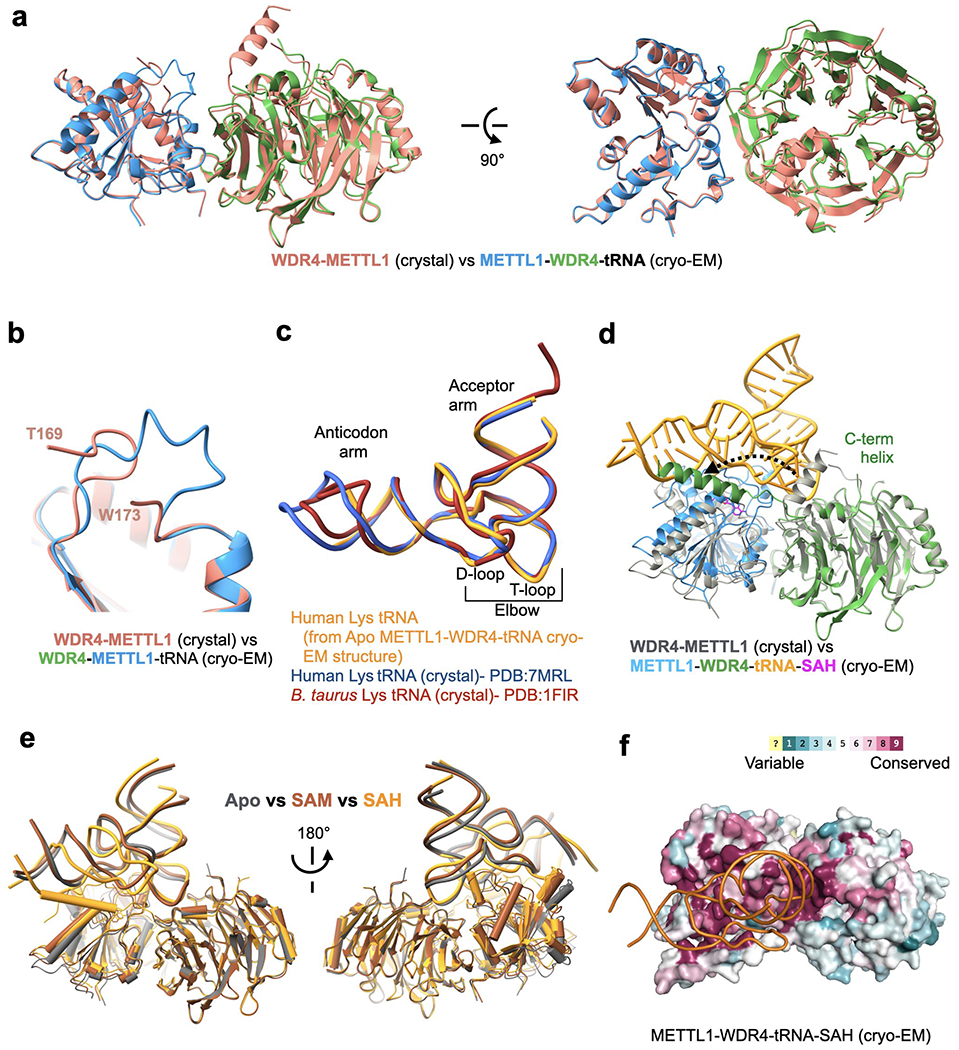
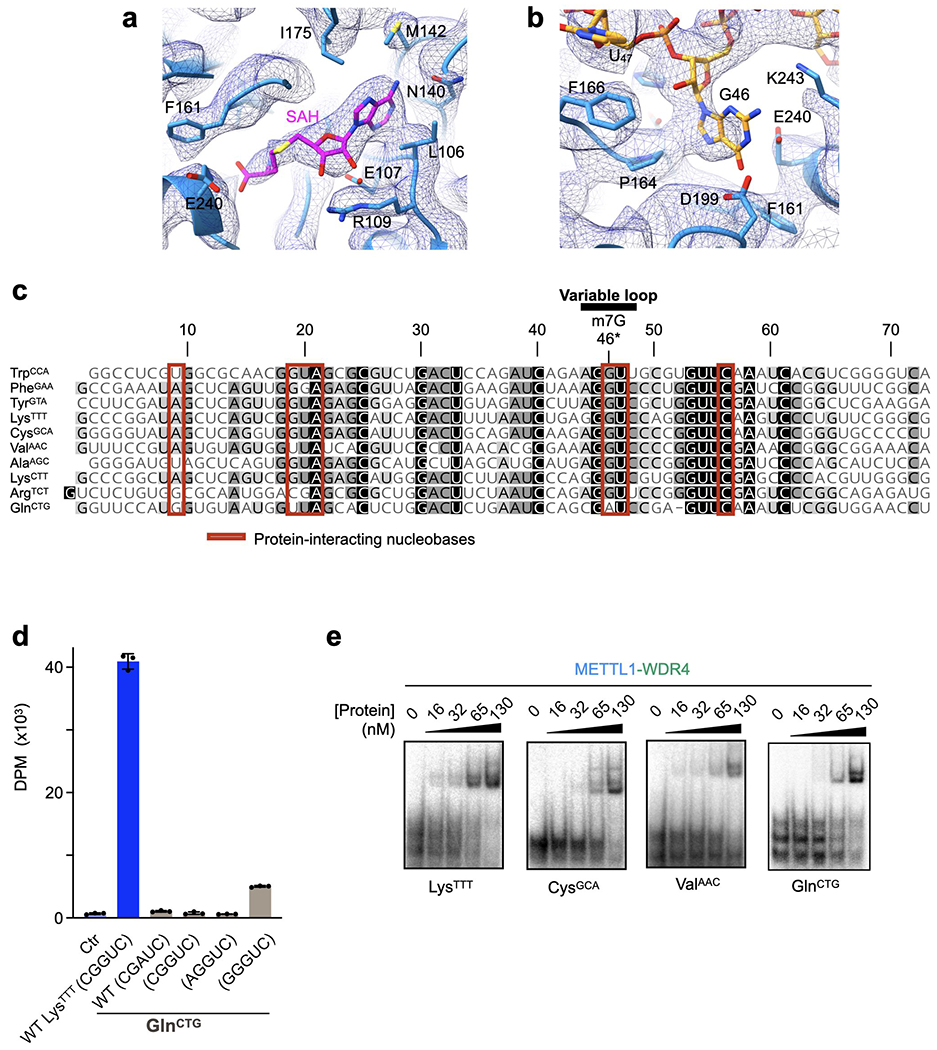
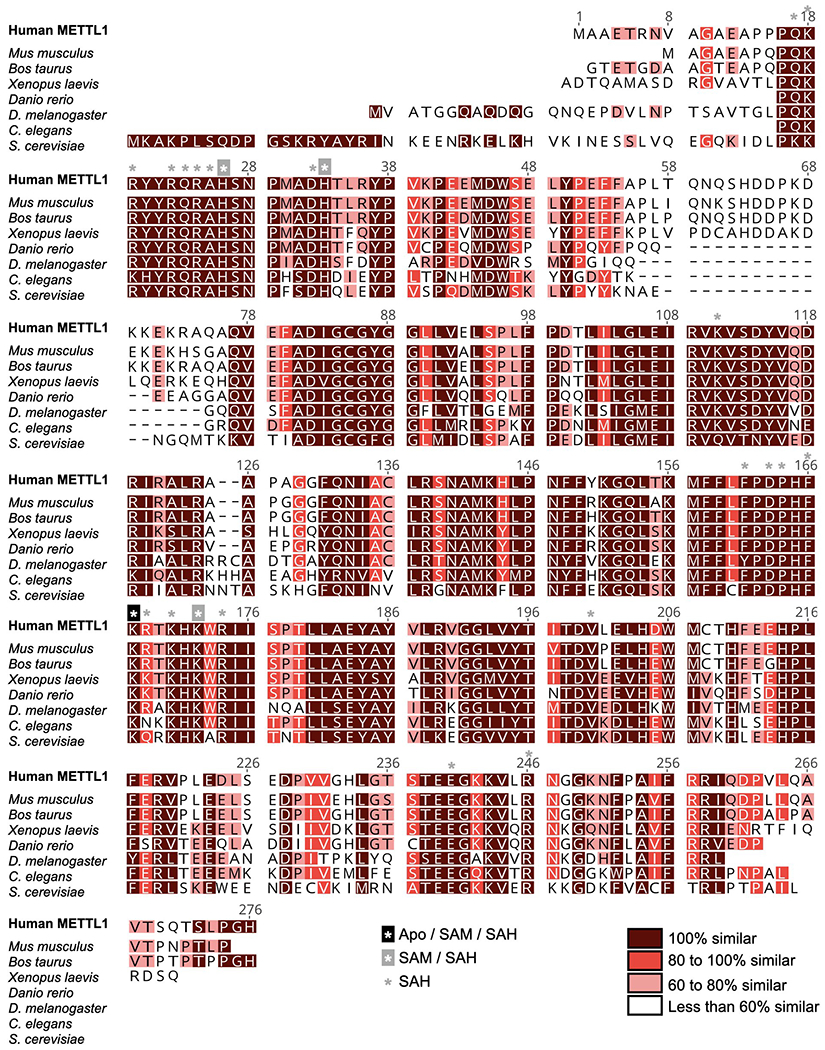
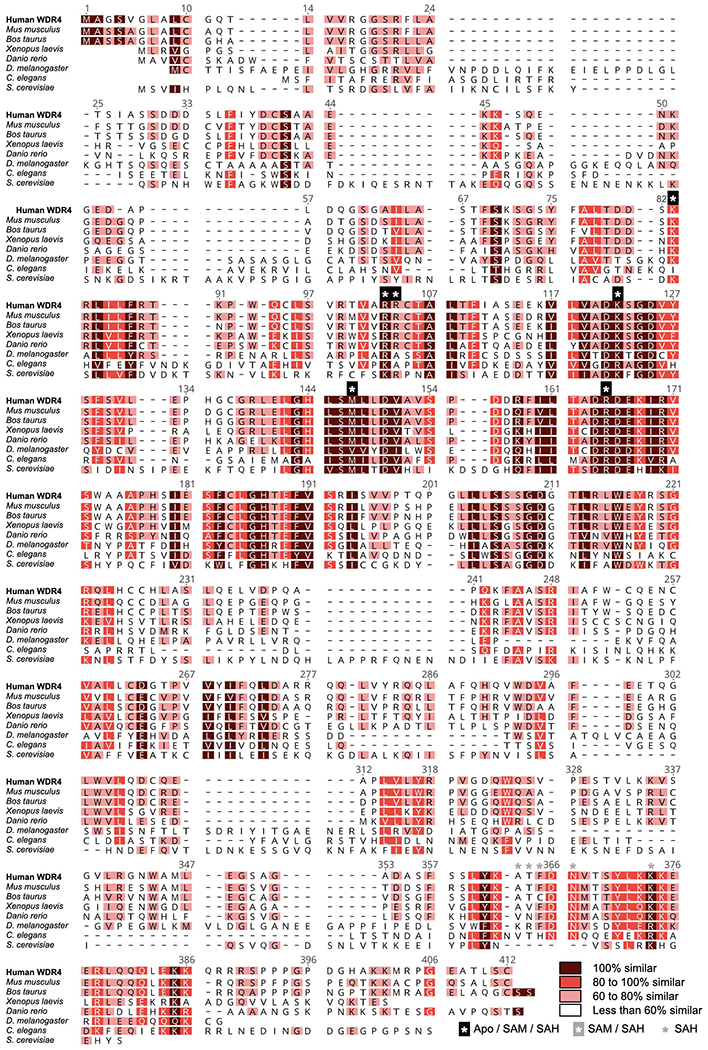
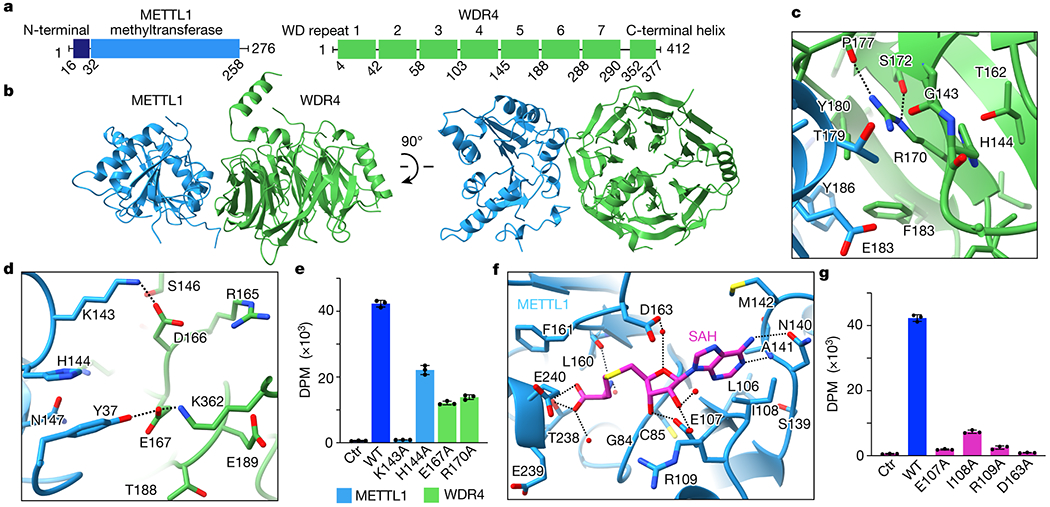
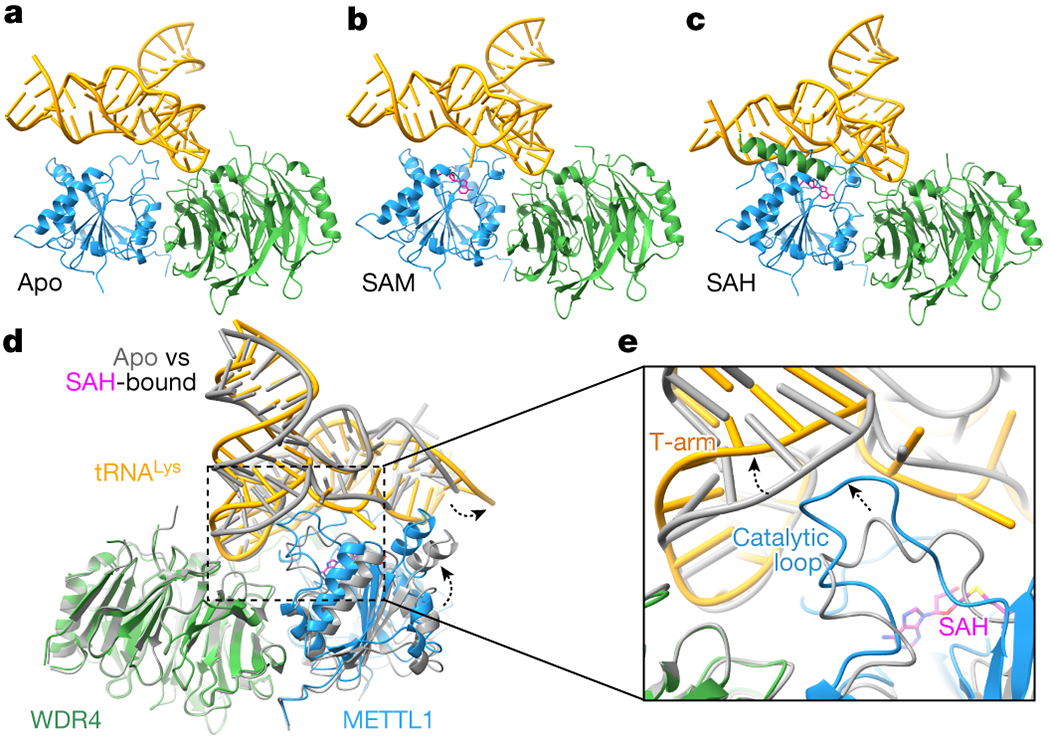
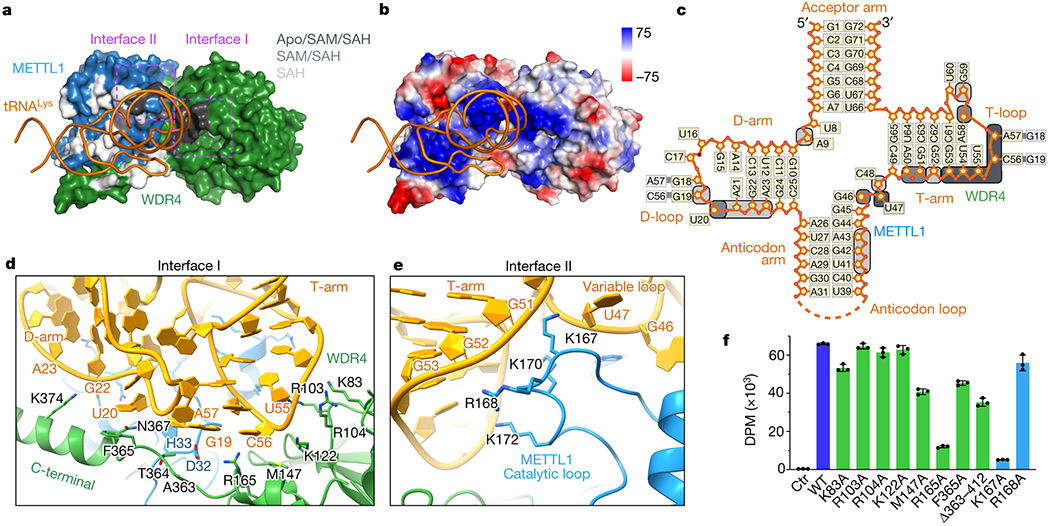
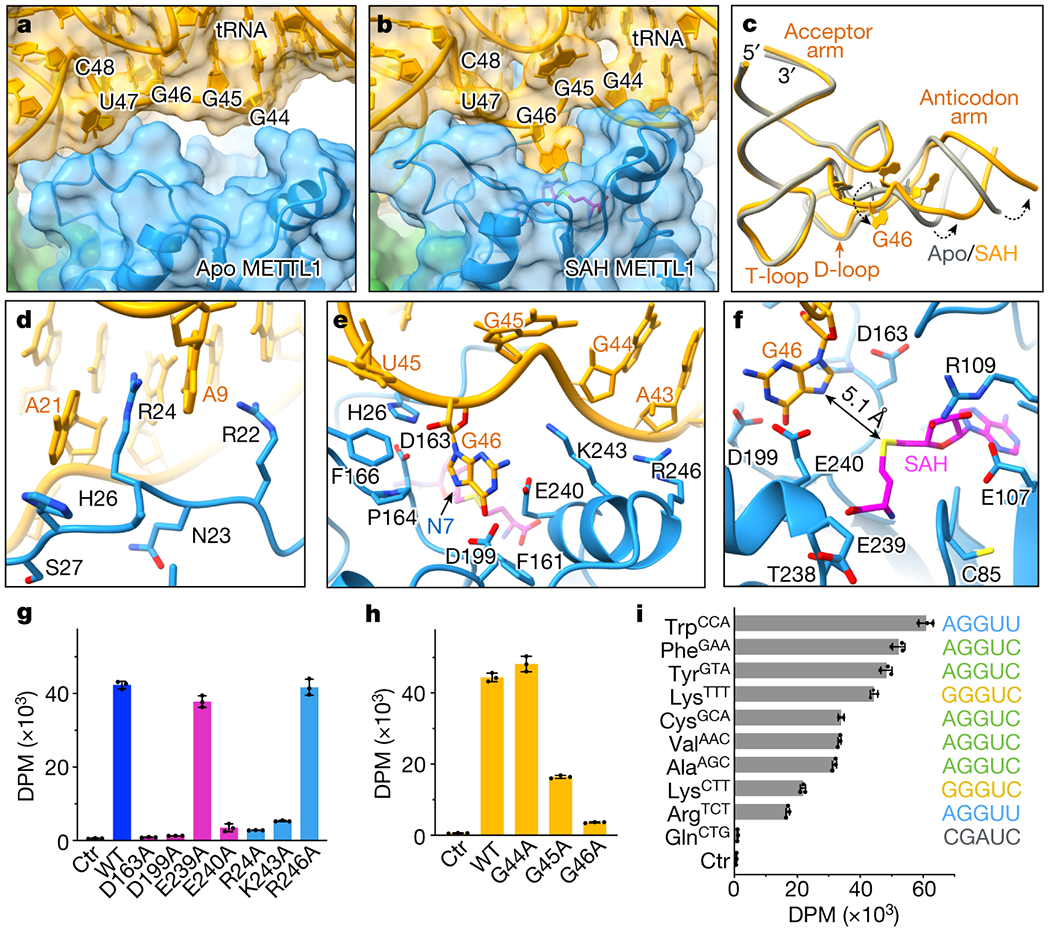
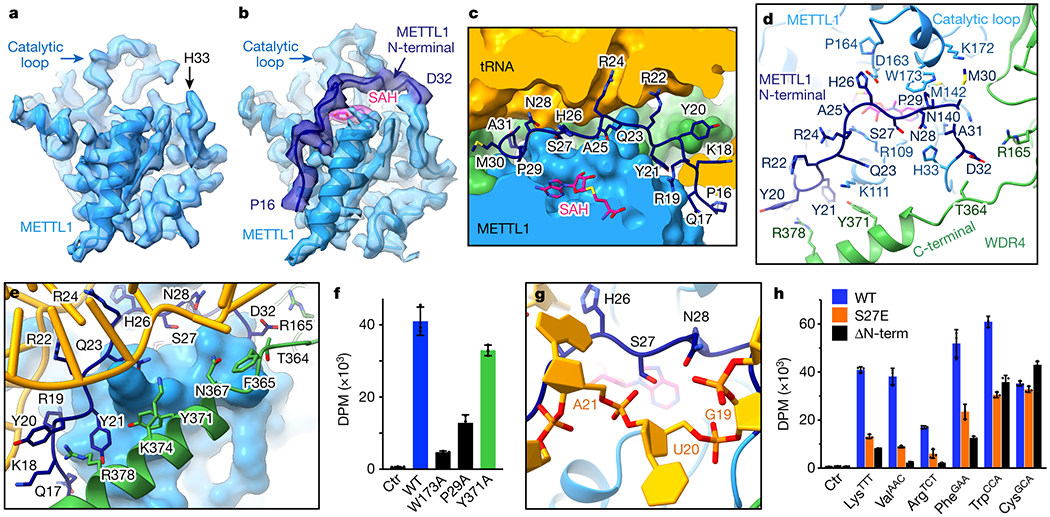
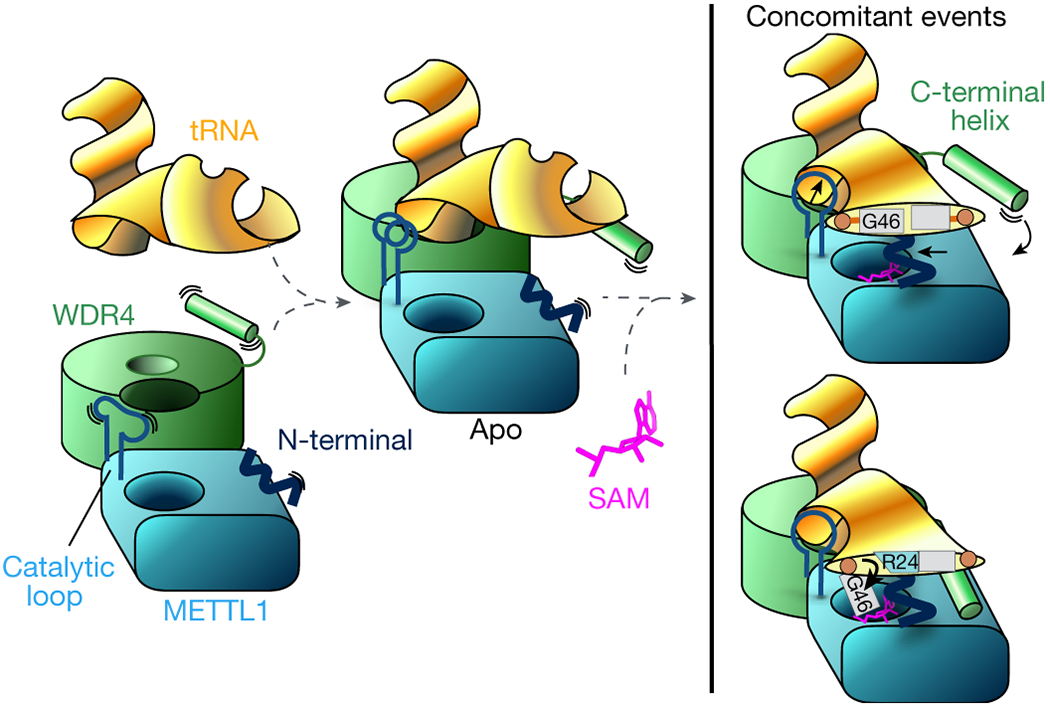
Specific, regulated modification of RNAs is important for proper gene expression1,2. tRNAs are rich with various chemical modifications that affect their stability and function3,4. 7-Methylguanosine (m7G) at tRNA position 46 is a conserved modification that modulates steady-state tRNA levels to affect cell growth5,6. The METTL1-WDR4 complex generates m7G46 in humans, and dysregulation of METTL1-WDR4 has been linked to brain malformation and multiple cancers7-22. Here we show how METTL1 and WDR4 cooperate to recognize RNA substrates and catalyse methylation. A crystal structure of METTL1-WDR4 and cryo-electron microscopy structures of METTL1-WDR4-tRNA show that the composite protein surface recognizes the tRNA elbow through shape complementarity. The cryo-electron microscopy structures of METTL1-WDR4-tRNA with S-adenosylmethionine or S-adenosylhomocysteine along with METTL1 crystal structures provide additional insights into the catalytic mechanism by revealing the active site in multiple states. The METTL1 N terminus couples cofactor binding with conformational changes in the tRNA, the catalytic loop and the WDR4 C terminus, acting as the switch to activate m7G methylation. Thus, our structural models explain how post-translational modifications of the METTL1 N terminus can regulate methylation. Together, our work elucidates the core and regulatory mechanisms underlying m7G modification by METTL1, providing the framework to understand its contribution to biology and disease.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures
References
References for Methods
-
- Studier FW Protein production by auto-induction in high density shaking cultures. Protein Expr Purif 41, 207–234 (2005). - PubMed
-
- Minor W, Cymborowski M, Otwinowski Z & Chruszcz M HKL-3000: the integration of data reduction and structure solution – from diffraction images to an initial model in minutes. Acta Crystallographica Section D: Biological Crystallography 62, 859–866 (2006). - PubMed
-
- Emsley P & Cowtan K Coot: model-building tools for molecular graphics. Acta Crystallogr D Biol Crystallogr 60, 2126–2132 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
