

Association between different sensory modalities based on concurrent time series data obtained by a collaborative reservoir computing model
- PMID: 36600034
- PMCID: PMC9813012
- DOI: 10.1038/s41598-023-27385-x
Association between different sensory modalities based on concurrent time series data obtained by a collaborative reservoir computing model
Abstract
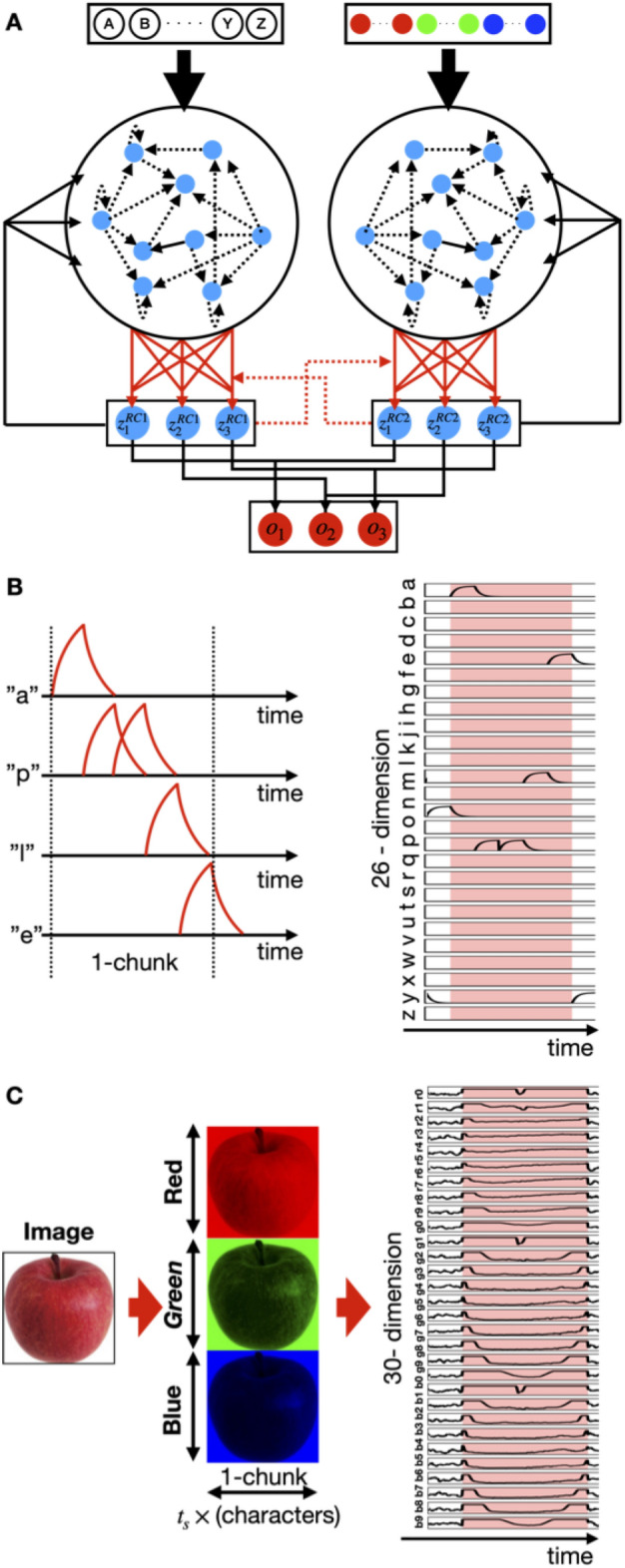
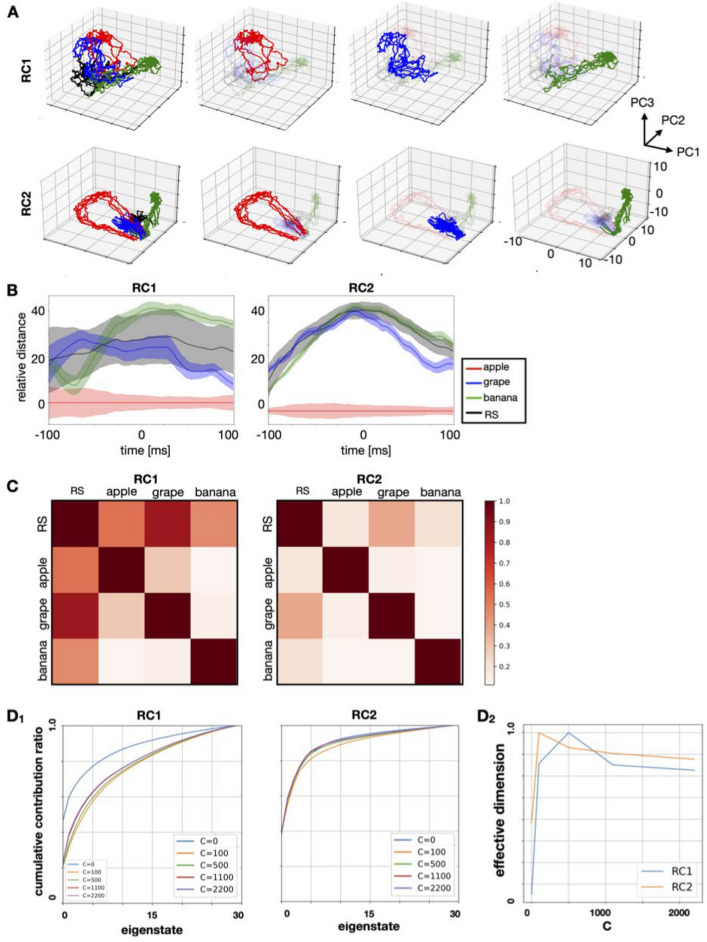
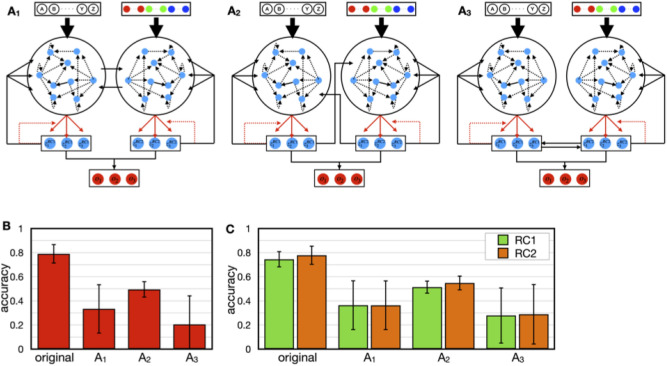
Humans perceive the external world by integrating information from different modalities, obtained through the sensory organs. However, the aforementioned mechanism is still unclear and has been a subject of widespread interest in the fields of psychology and brain science. A model using two reservoir computing systems, i.e., a type of recurrent neural network trained to mimic each other's output, can detect stimulus patterns that repeatedly appear in a time series signal. We applied this model for identifying specific patterns that co-occur between information from different modalities. The model was self-organized by specific fluctuation patterns that co-occurred between different modalities, and could detect each fluctuation pattern. Additionally, similarly to the case where perception is influenced by synchronous/asynchronous presentation of multimodal stimuli, the model failed to work correctly for signals that did not co-occur with corresponding fluctuation patterns. Recent experimental studies have suggested that direct interaction between different sensory systems is important for multisensory integration, in addition to top-down control from higher brain regions such as the association cortex. Because several patterns of interaction between sensory modules can be incorporated into the employed model, we were able to compare the performance between them; the original version of the employed model incorporated such an interaction as the teaching signals for learning. The performance of the original and alternative models was evaluated, and the original model was found to perform the best. Thus, we demonstrated that feedback of the outputs of appropriately learned sensory modules performed the best when compared to the other examined patterns of interaction. The proposed model incorporated information encoded by the dynamic state of the neural population and the interactions between different sensory modules, both of which were based on recent experimental observations; this allowed us to study the influence of the temporal relationship and frequency of occurrence of multisensory signals on sensory integration, as well as the nature of interaction between different sensory signals.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
