

Clonally expanded HIV-1 proviruses with 5'-leader defects can give rise to nonsuppressible residual viremia
- PMID: 36602866
- PMCID: PMC10014112
- DOI: 10.1172/JCI165245
Clonally expanded HIV-1 proviruses with 5'-leader defects can give rise to nonsuppressible residual viremia
Abstract
BackgroundAntiretroviral therapy (ART) halts HIV-1 replication, decreasing viremia to below the detection limit of clinical assays. However, some individuals experience persistent nonsuppressible viremia (NSV) originating from CD4+ T cell clones carrying infectious proviruses. Defective proviruses represent over 90% of all proviruses persisting during ART and can express viral genes, but whether they can cause NSV and complicate ART management is unknown.MethodsWe undertook an in-depth characterization of proviruses causing NSV in 4 study participants with optimal adherence and no drug resistance. We investigated the impact of the observed defects on 5'-leader RNA properties, virus infectivity, and gene expression. Integration-site specific assays were used to track these proviruses over time and among cell subsets.ResultsClones carrying proviruses with 5'-leader defects can cause persistent NSV up to approximately 103 copies/mL. These proviruses had small, often identical deletions or point mutations involving the major splicing donor (MSD) site and showed partially reduced RNA dimerization and nucleocapsid binding. Nevertheless, they were inducible and produced noninfectious virions containing viral RNA, but lacking envelope.ConclusionThese findings show that proviruses with 5'-leader defects in CD4+ T cell clones can give rise to NSV, affecting clinical care. Sequencing of the 5'-leader can help in understanding failure to completely suppress viremia.FundingOffice of the NIH Director and National Institute of Dental and Craniofacial Research, NIH; Howard Hughes Medical Institute; Johns Hopkins University Center for AIDS Research; National Institute for Allergy and Infectious Diseases (NIAID), NIH, to the PAVE, BEAT-HIV, and DARE Martin Delaney collaboratories.
Keywords: AIDS/HIV; T cells.
Figures
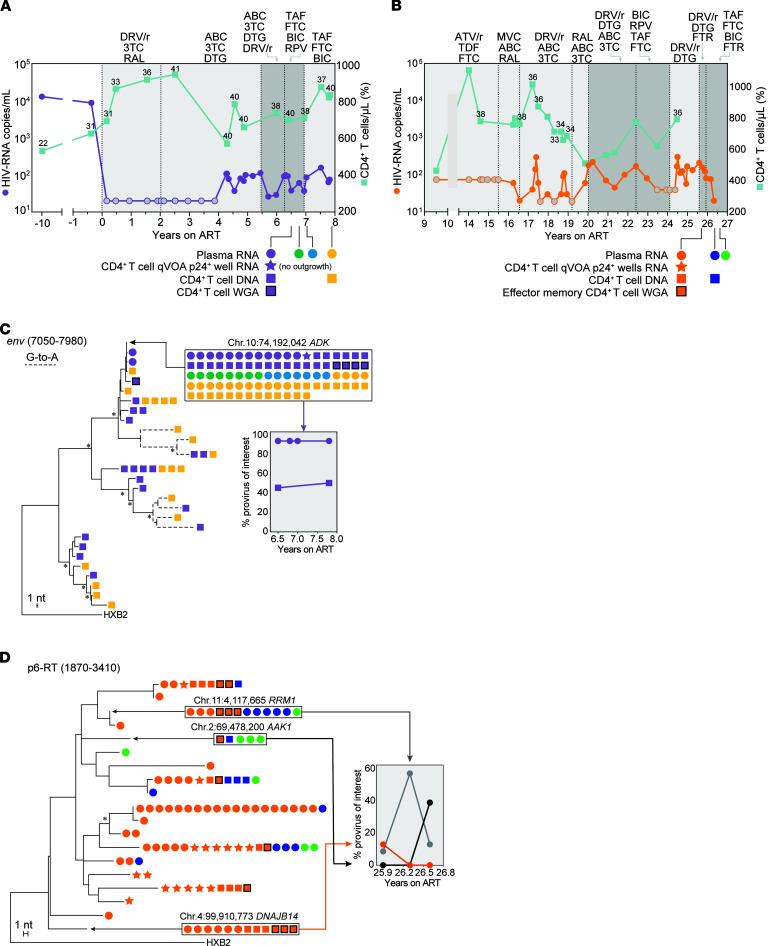
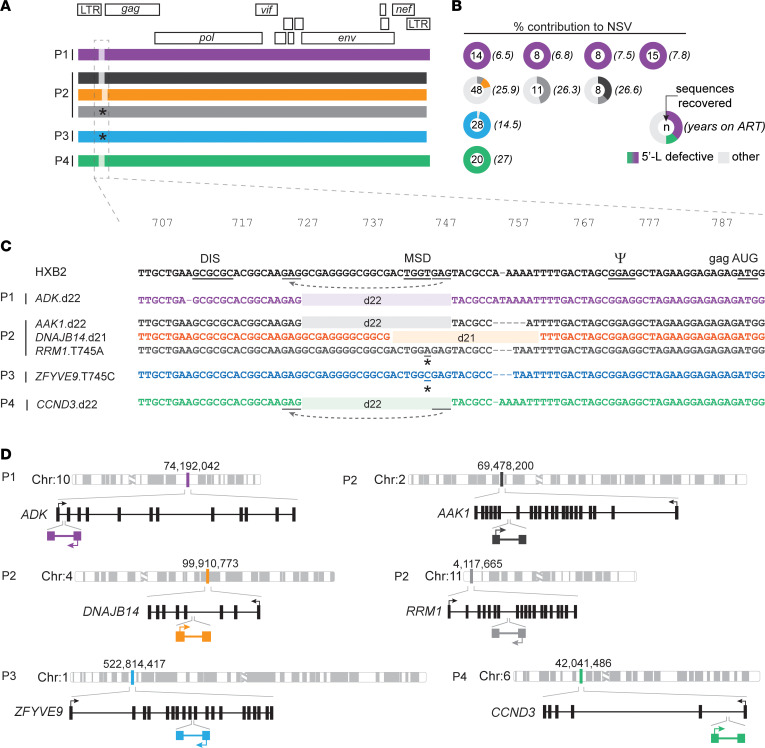





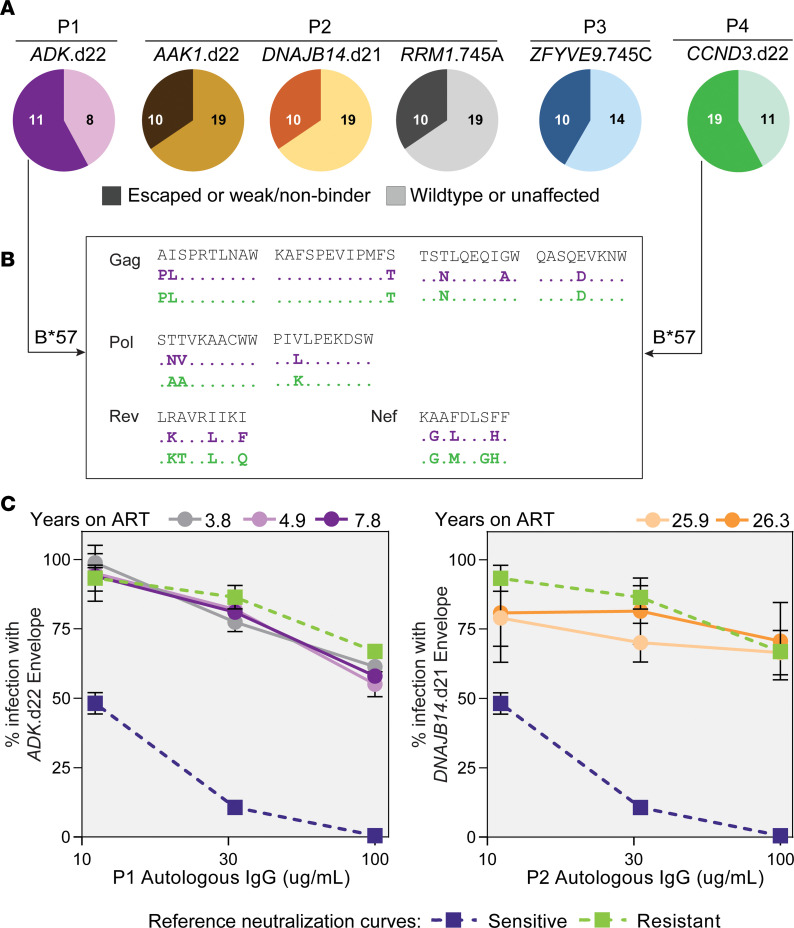
Comment in
- Nonsuppressible viremia during HIV-1 therapy meets molecular virology doi: 10.1172/JCI167925
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
