

Muscarinic Acetylcholine Receptors Modulate HCN Channel Properties in Vestibular Ganglion Neurons
- PMID: 36604171
- PMCID: PMC9908319
- DOI: 10.1523/JNEUROSCI.2552-21.2022
Muscarinic Acetylcholine Receptors Modulate HCN Channel Properties in Vestibular Ganglion Neurons
Abstract
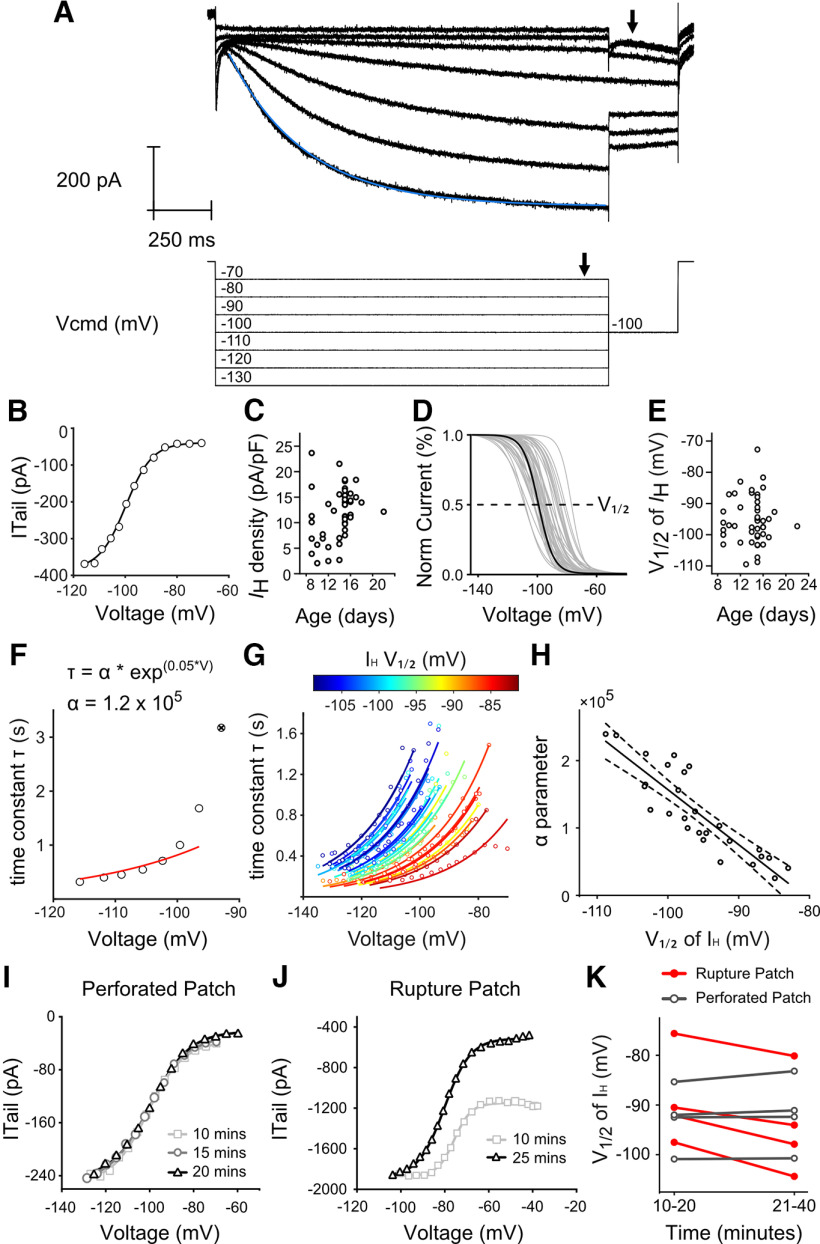
Efferent modulation of vestibular afferent excitability is linked to muscarinic signaling cascades that close low-voltage-gated potassium channels (i.e., KCNQ). Here, we show that muscarinic signaling cascades also depolarize the activation range of hyperpolarization-activated cyclic-nucleotide gated (HCN) channels. We compared the voltage activation range and kinetics of HCN channels and induced firing patterns before and after administering the muscarinic acetylcholine receptor (mAChR) agonist oxotremorine-M (Oxo-M) in dissociated vestibular ganglion neurons (VGNs) from rats of either sex using perforated whole-cell patch-clamp methods. Oxo-M depolarized HCN channels' half-activation voltage (V 1/2) and sped up the rate of activation near resting potential twofold. HCN channels in large-diameter and/or transient firing VGN (putative cell bodies of irregular firing neuron from central epithelial zones) had relatively depolarized V 1/2 in control solution and were less sensitive to mAChR activation than those found in small-diameter VGN with sustained firing patterns (putatively belonging to regular firing afferents). The impact of mAChR on HCN channels is not a direct consequence of closing KCNQ channels since pretreating the cells with Linopirdine, a KCNQ channel blocker, did not prevent HCN channel depolarization by Oxo-M. Efferent signaling promoted ion channel configurations that were favorable to highly regular spiking in some VGN, but not others. This is consistent with previous observations that low-voltage gated potassium currents in VGN are conducted by mAChR agonist-sensitive and -insensitive channels. Connecting efferent signaling to HCN channels is significant because of the channel's impact on spike-timing regularity and nonchemical transmission between Type I hair cells and vestibular afferents.SIGNIFICANCE STATEMENT Vestibular afferents express a diverse complement of ion channels. In vitro studies identified low-voltage activated potassium channels and hyperpolarization-activated cyclic-nucleotide gated (HCN) channels as crucial for shaping the timing and sensitivity of afferent responses. Moreover, a network of acetylcholine-releasing efferent neurons controls afferent excitability by closing a subgroup of low-voltage activated potassium channels on the afferent neuron. This work shows that these efferent signaling cascades also enhance the activation of HCN channels by depolarizing their voltage activation range. The size of this effect varies depending on the endogenous properties of the HCN channel and on cell type (as determined by discharge patterns and cell size). Simultaneously controlling two ion-channel groups gives the vestibular efferent system exquisite control over afferent neuron activity.
Keywords: HCN channels; KCNQ channels; efferent neurons; modulation; muscarinic acetylcholine receptors; vestibular ganglion.
Copyright © 2023 the authors.
Figures






Similar articles
-
Enhanced Activation of HCN Channels Reduces Excitability and Spike-Timing Regularity in Maturing Vestibular Afferent Neurons.J Neurosci. 2019 Apr 10;39(15):2860-2876. doi: 10.1523/JNEUROSCI.1811-18.2019. Epub 2019 Jan 29. J Neurosci. 2019. PMID: 30696730 Free PMC article.
-
Muscarinic Acetylcholine Receptors and M-Currents Underlie Efferent-Mediated Slow Excitation in Calyx-Bearing Vestibular Afferents.J Neurosci. 2017 Feb 15;37(7):1873-1887. doi: 10.1523/JNEUROSCI.2322-16.2017. Epub 2017 Jan 16. J Neurosci. 2017. PMID: 28093476 Free PMC article.
-
Functional impact of the hyperpolarization-activated current on the excitability of myelinated A-type vagal afferent neurons in the rat.Clin Exp Pharmacol Physiol. 2010 Aug;37(8):852-61. doi: 10.1111/j.1440-1681.2010.05396.x. Epub 2010 Apr 26. Clin Exp Pharmacol Physiol. 2010. PMID: 20456426 Free PMC article.
-
Neural KCNQ (Kv7) channels.Br J Pharmacol. 2009 Apr;156(8):1185-95. doi: 10.1111/j.1476-5381.2009.00111.x. Epub 2009 Mar 9. Br J Pharmacol. 2009. PMID: 19298256 Free PMC article. Review.
-
Hyperpolarization-activated cyclic nucleotide-gated (HCN) channels and pain.Curr Pharm Des. 2009;15(15):1767-72. doi: 10.2174/138161209788186281. Curr Pharm Des. 2009. PMID: 19442189 Review.
Cited by
-
Isolating and Culturing Vestibular and Spiral Ganglion Somata from Neonatal Rodents for Patch-Clamp Recordings.J Vis Exp. 2023 Apr 21;(194):10.3791/64908. doi: 10.3791/64908. J Vis Exp. 2023. PMID: 37154564 Free PMC article.
-
Expression of hyperpolarization-activated current (Ih) in zonally defined vestibular calyx terminals of the crista.J Neurophysiol. 2023 Jun 1;129(6):1468-1481. doi: 10.1152/jn.00135.2023. Epub 2023 May 17. J Neurophysiol. 2023. PMID: 37198134 Free PMC article.
-
Muscarinic receptor activation preferentially inhibits rebound in vulnerable dopaminergic neurons.bioRxiv [Preprint]. 2024 Jul 31:2024.07.30.605819. doi: 10.1101/2024.07.30.605819. bioRxiv. 2024. Update in: J Neurosci. 2025 Apr 16;45(16):e1443242025. doi: 10.1523/JNEUROSCI.1443-24.2025. PMID: 39131326 Free PMC article. Updated. Preprint.
-
Muscarinic Receptor Activation Preferentially Inhibits Rebound in Vulnerable Dopaminergic Neurons.J Neurosci. 2025 Apr 16;45(16):e1443242025. doi: 10.1523/JNEUROSCI.1443-24.2025. J Neurosci. 2025. PMID: 40000233
-
Contribution of membrane-associated oscillators to biological timing at different timescales.Front Physiol. 2024 Jan 9;14:1243455. doi: 10.3389/fphys.2023.1243455. eCollection 2023. Front Physiol. 2024. PMID: 38264332 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources