

Enteroendocrine Cells Protect the Stem Cell Niche by Regulating Crypt Metabolism in Response to Nutrients
- PMID: 36608902
- PMCID: PMC10140799
- DOI: 10.1016/j.jcmgh.2022.12.016
Enteroendocrine Cells Protect the Stem Cell Niche by Regulating Crypt Metabolism in Response to Nutrients
Abstract
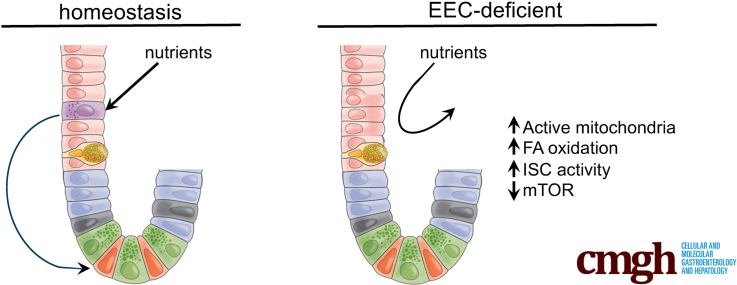
Background & aims: The intestinal stem cell niche is exquisitely sensitive to changes in diet, with high-fat diet, caloric restriction, and fasting resulting in altered crypt metabolism and intestinal stem cell function. Unlike cells on the villus, cells in the crypt are not immediately exposed to the dynamically changing contents of the lumen. We hypothesized that enteroendocrine cells (EECs), which sense environmental cues and in response release hormones and metabolites, are essential for relaying the luminal and nutritional status of the animal to cells deep in the crypt.
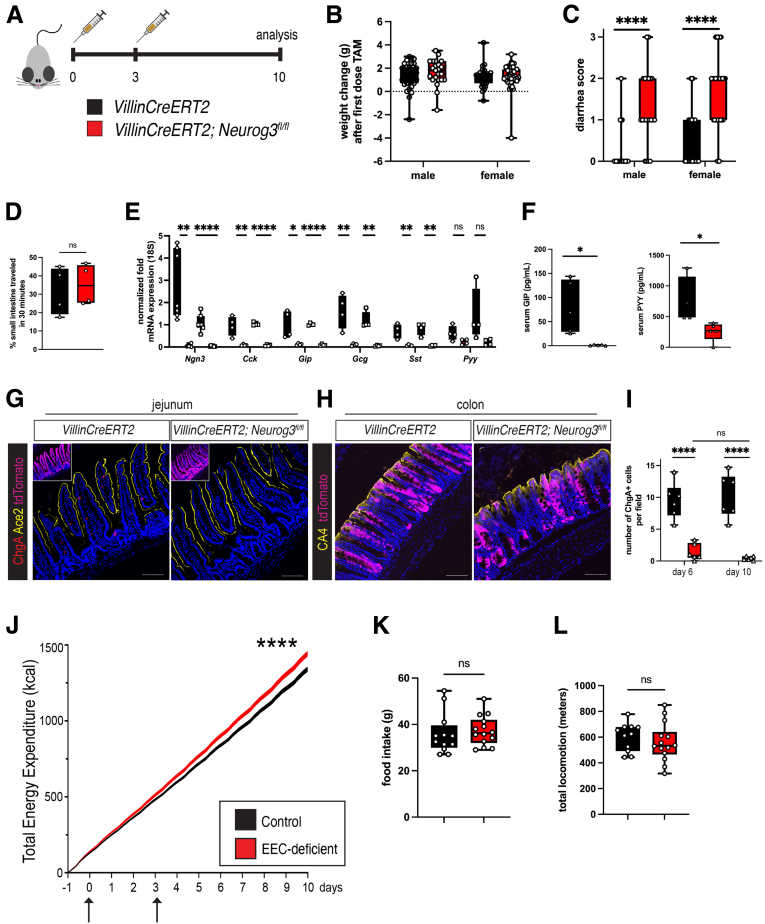
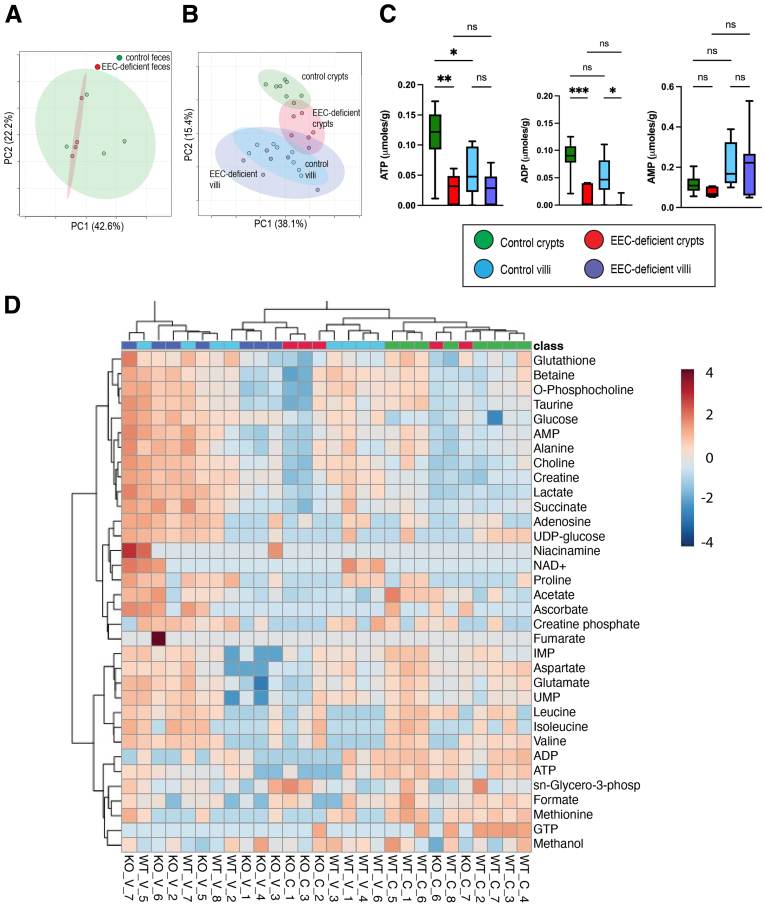
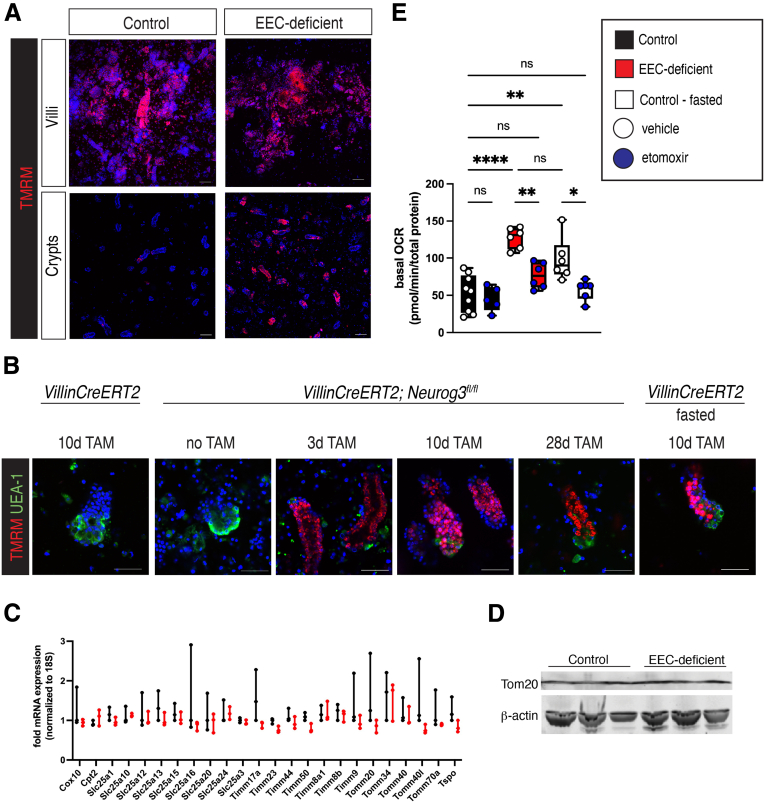
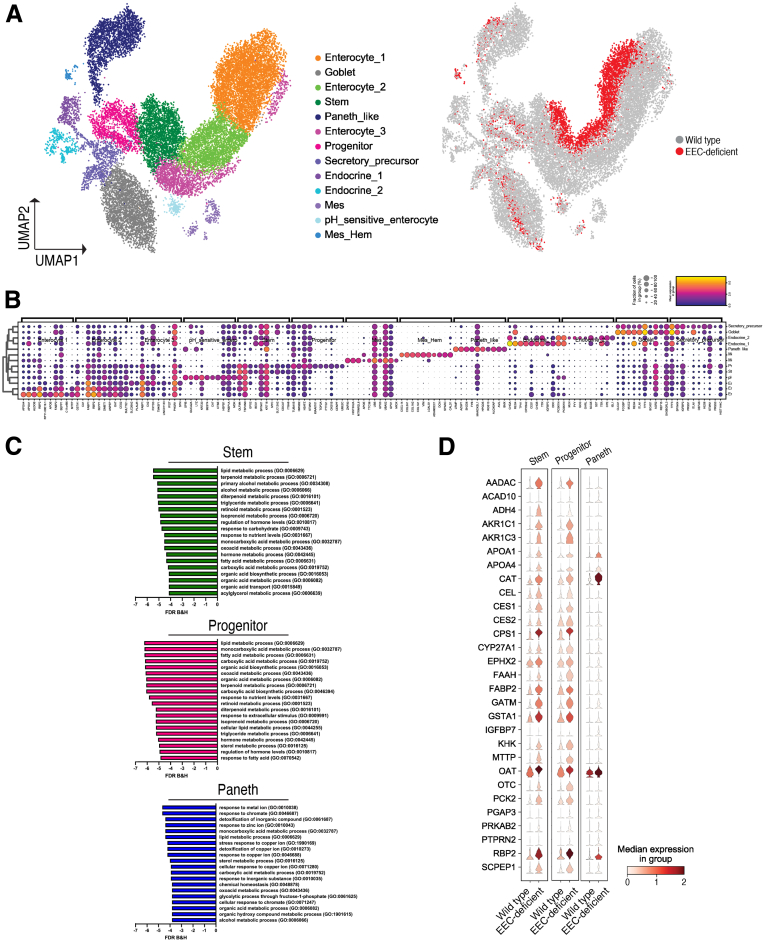
Methods: We used the tamoxifen-inducible VillinCreERT2 mouse model to deplete EECs (Neurog3fl/fl) from adult intestinal epithelium and we generated human intestinal organoids from wild-type and NEUROGENIN 3 (NEUROG3)-null human pluripotent stem cells. We used indirect calorimetry, 1H-Nuclear Magnetic Resonance (NMR) metabolomics, mitochondrial live imaging, and the Seahorse bioanalyzer (Agilent Technologies) to assess metabolism. Intestinal stem cell activity was measured by proliferation and enteroid-forming capacity. Transcriptional changes were assessed using 10x Genomics single-cell sequencing.
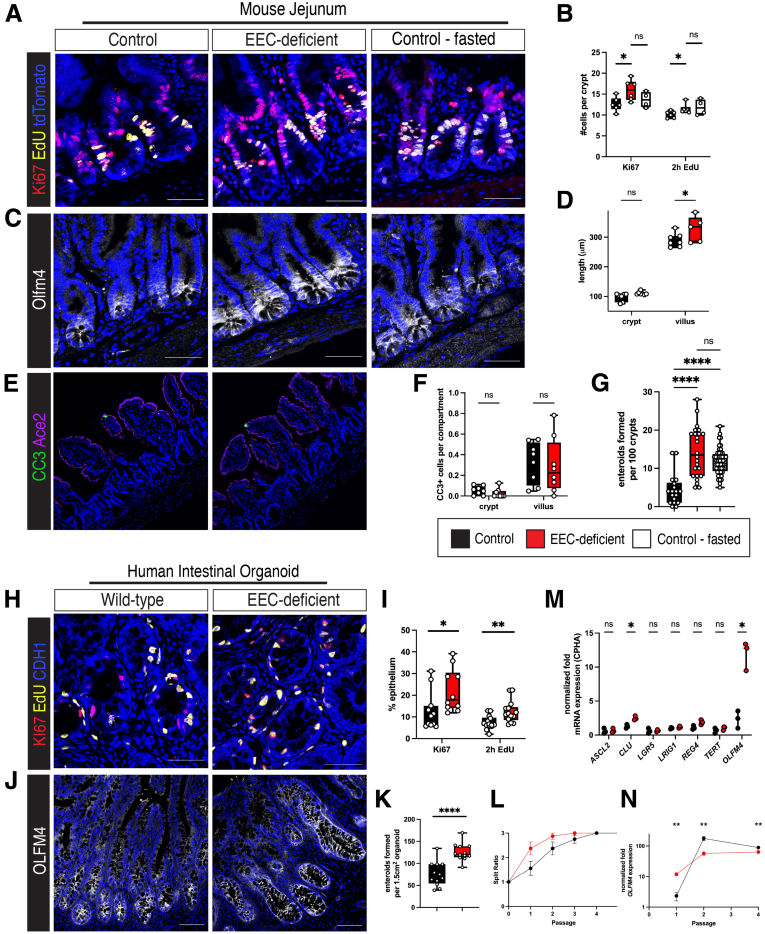
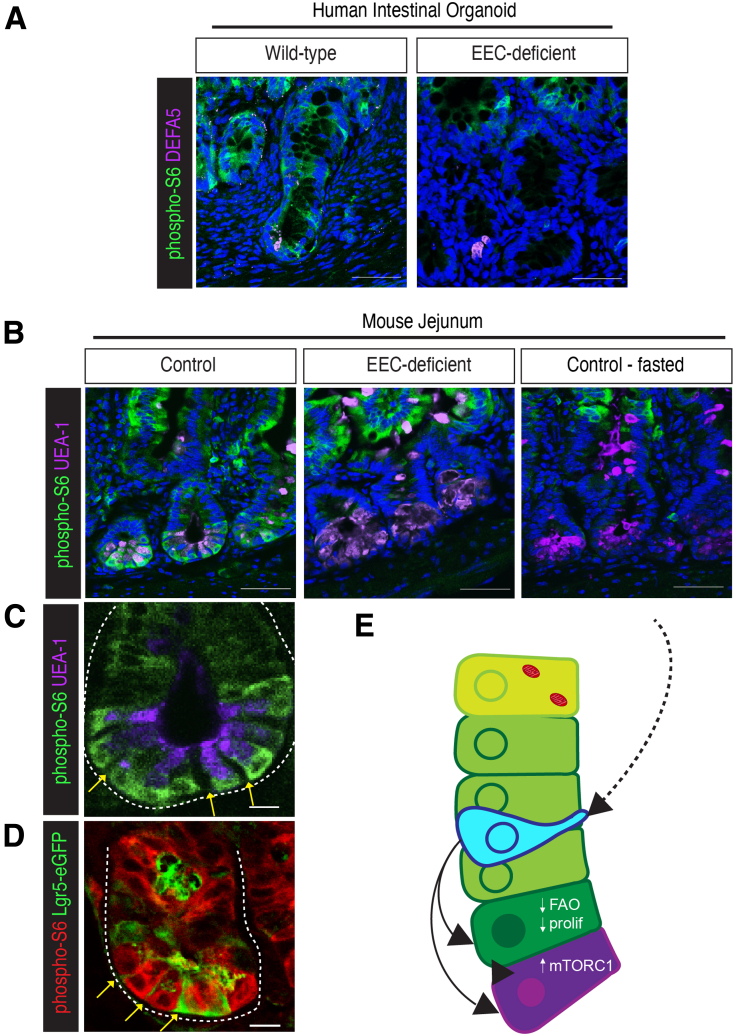
Results: Loss of EECs resulted in increased energy expenditure in mice, an abundance of active mitochondria, and a shift of crypt metabolism to fatty acid oxidation. Crypts from mouse and human intestinal organoids lacking EECs displayed increased intestinal stem cell activity and failed to activate phosphorylation of downstream target S6 kinase ribosomal protein, a marker for activity of the master metabolic regulator mammalian target of rapamycin (mTOR). These phenotypes were similar to those observed when control mice were deprived of nutrients.
Conclusions: EECs are essential regulators of crypt metabolism. Depletion of EECs recapitulated a fasting metabolic phenotype despite normal levels of ingested nutrients. These data suggest that EECs are required to relay nutritional information to the stem cell niche and are essential regulators of intestinal metabolism.
Keywords: Enteroendocrine Cells; Intestinal Metabolism; Intestinal Stem Cell; Mitochondria.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
A New Role for Endocrine Cells in the Intestinal Crypt.Cell Mol Gastroenterol Hepatol. 2023;15(6):1525-1526. doi: 10.1016/j.jcmgh.2023.01.012. Epub 2023 Feb 17. Cell Mol Gastroenterol Hepatol. 2023. PMID: 36805429 Free PMC article. No abstract available.
-
Nutrient Sensing and Lipid Handling by Enteroendocrine Cells in Adult Mice.Cell Mol Gastroenterol Hepatol. 2023;15(6):1535-1536. doi: 10.1016/j.jcmgh.2023.03.006. Epub 2023 Apr 6. Cell Mol Gastroenterol Hepatol. 2023. PMID: 37031707 Free PMC article. No abstract available.
References
-
- Duée P.-H., Darcy-Vrillon B., Blachier F., Morel M.-T. Fuel selection in intestinal cells. Proc Nutr Soc. 1995;54:83–94. - PubMed
-
- Beumer J., Clevers H. Cell fate specification and differentiation in the adult mammalian intestine. Nat Rev Mol Cell Biol. 2021;22:39–53. - PubMed
-
- Gribble F.M., Reimann F. Enteroendocrine cells: chemosensors in the intestinal epithelium. Annu Rev Physiol. 2016;78:277–299. - PubMed
-
- Wang J., Cortina G., Wu S.V., et al. Mutant neurogenin-3 in congenital malabsorptive diarrhea. N Engl J Med. 2006;355:270–280. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
