

Optic flow based spatial vision in insects
- PMID: 36609568
- PMCID: PMC10354154
- DOI: 10.1007/s00359-022-01610-w
Optic flow based spatial vision in insects
Abstract
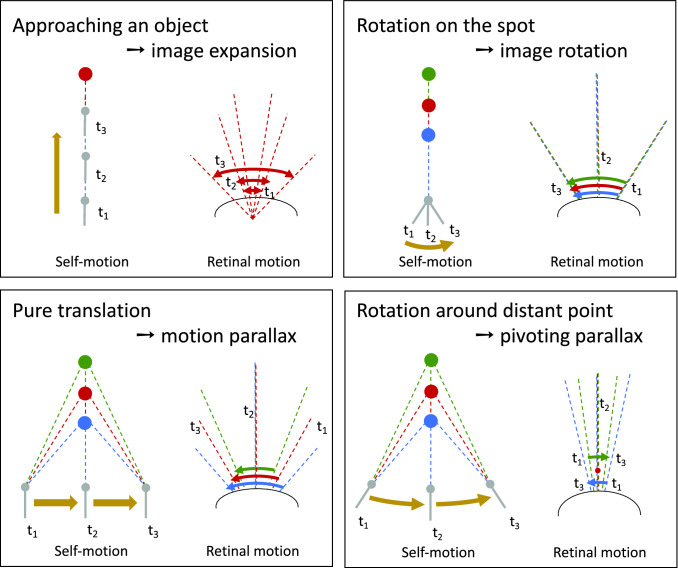
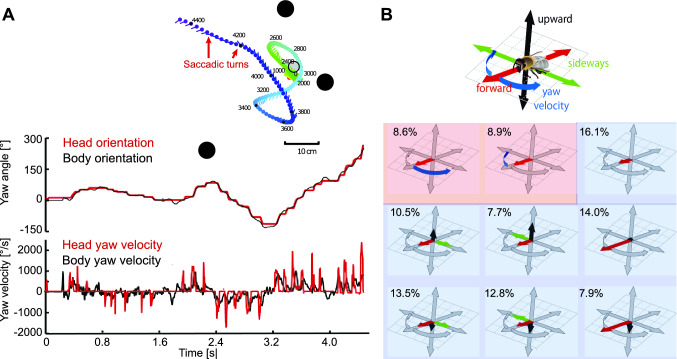
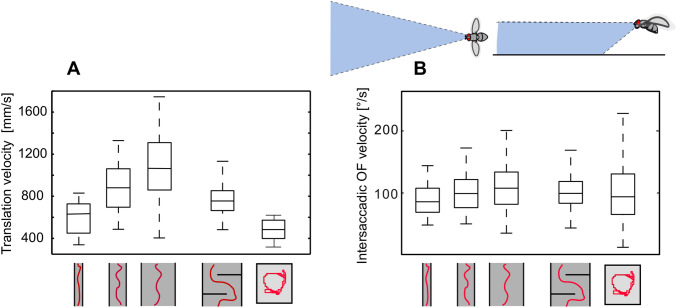
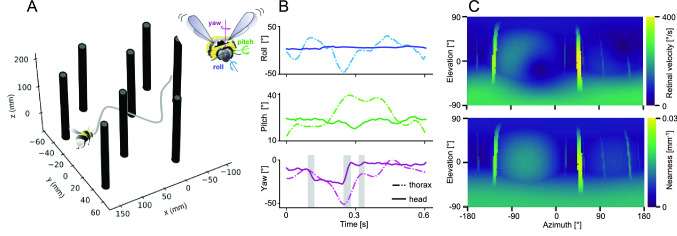
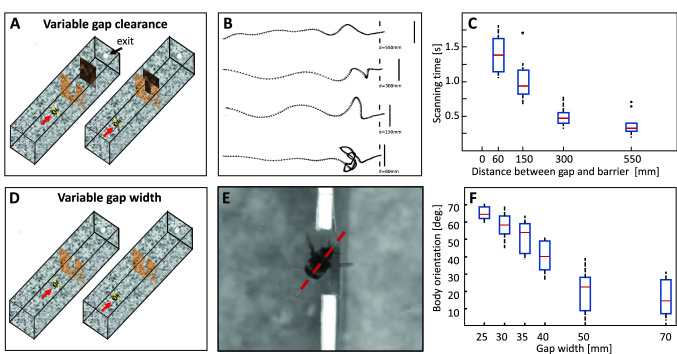
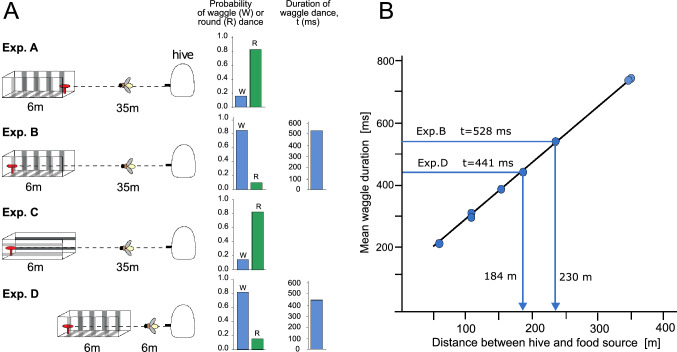
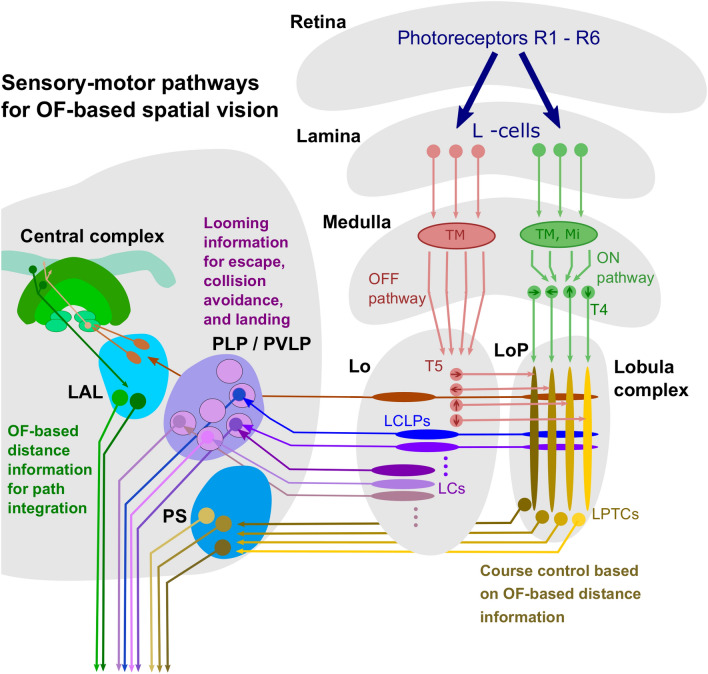
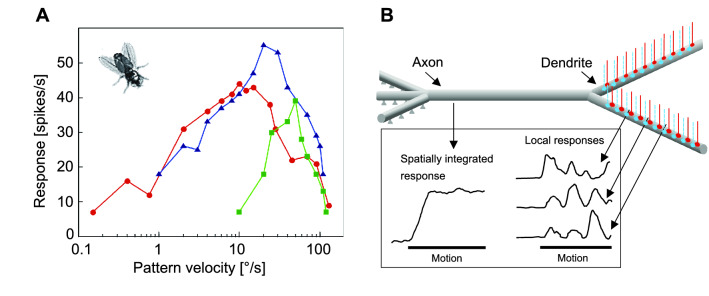
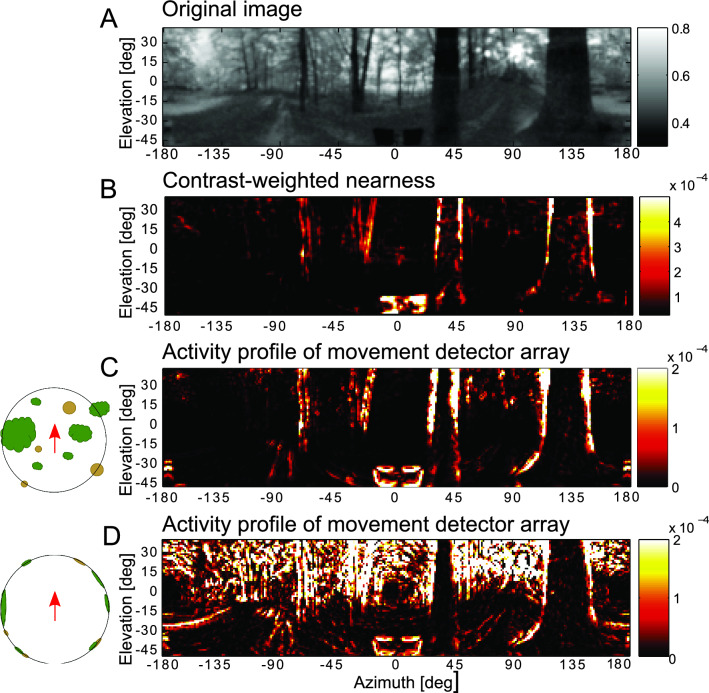
The optic flow, i.e., the displacement of retinal images of objects in the environment induced by self-motion, is an important source of spatial information, especially for fast-flying insects. Spatial information over a wide range of distances, from the animal's immediate surroundings over several hundred metres to kilometres, is necessary for mediating behaviours, such as landing manoeuvres, collision avoidance in spatially complex environments, learning environmental object constellations and path integration in spatial navigation. To facilitate the processing of spatial information, the complexity of the optic flow is often reduced by active vision strategies. These result in translations and rotations being largely separated by a saccadic flight and gaze mode. Only the translational components of the optic flow contain spatial information. In the first step of optic flow processing, an array of local motion detectors provides a retinotopic spatial proximity map of the environment. This local motion information is then processed in parallel neural pathways in a task-specific manner and used to control the different components of spatial behaviour. A particular challenge here is that the distance information extracted from the optic flow does not represent the distances unambiguously, but these are scaled by the animal's speed of locomotion. Possible ways of coping with this ambiguity are discussed.
Keywords: Behavioural control; Motion detection; Optic flow; Spatial vision.
© 2023. The Author(s).
Conflict of interest statement
The author declares that he has no conflict of interest.
Figures
Similar articles
-
Path integration and optic flow in flying insects: a review of current evidence.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025 May;211(3):375-401. doi: 10.1007/s00359-025-01734-9. Epub 2025 Mar 7. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025. PMID: 40053081 Free PMC article. Review.
-
Local motion adaptation enhances the representation of spatial structure at EMD arrays.PLoS Comput Biol. 2017 Dec 27;13(12):e1005919. doi: 10.1371/journal.pcbi.1005919. eCollection 2017 Dec. PLoS Comput Biol. 2017. PMID: 29281631 Free PMC article.
-
A Bio-inspired Collision Avoidance Model Based on Spatial Information Derived from Motion Detectors Leads to Common Routes.PLoS Comput Biol. 2015 Nov 19;11(11):e1004339. doi: 10.1371/journal.pcbi.1004339. eCollection 2015 Nov. PLoS Comput Biol. 2015. PMID: 26583771 Free PMC article.
-
Optic flow-based collision-free strategies: From insects to robots.Arthropod Struct Dev. 2017 Sep;46(5):703-717. doi: 10.1016/j.asd.2017.06.003. Epub 2017 Jul 11. Arthropod Struct Dev. 2017. PMID: 28655645 Review.
-
Optic flow-based course control in insects.Curr Opin Neurobiol. 2020 Feb;60:21-27. doi: 10.1016/j.conb.2019.10.007. Epub 2019 Dec 3. Curr Opin Neurobiol. 2020. PMID: 31810007 Review.
Cited by
-
Visual physiology of Australian stingless bees.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025 Jul;211(4):435-444. doi: 10.1007/s00359-025-01740-x. Epub 2025 May 20. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025. PMID: 40392294 Free PMC article.
-
trajPredRNN+: A new approach for precipitation nowcasting with weather radar echo images based on deep learning.Heliyon. 2024 Aug 10;10(18):e36134. doi: 10.1016/j.heliyon.2024.e36134. eCollection 2024 Sep 30. Heliyon. 2024. PMID: 39309946 Free PMC article.
-
Bumblebees locate goals in 3D with absolute height estimation from ventral optic flow.J Exp Biol. 2025 Jun 1;228(11):jeb249763. doi: 10.1242/jeb.249763. Epub 2025 Jun 10. J Exp Biol. 2025. PMID: 40353641 Free PMC article.
-
Path integration and optic flow in flying insects: a review of current evidence.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025 May;211(3):375-401. doi: 10.1007/s00359-025-01734-9. Epub 2025 Mar 7. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2025. PMID: 40053081 Free PMC article. Review.
-
Integration of parallel pathways for flight control in a hawkmoth reflects prevalence and relevance of natural visual cues.Elife. 2025 Jul 17;14:RP104118. doi: 10.7554/eLife.104118. Elife. 2025. PMID: 40673488 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
